Om efteråret har fårekyllinger generelt en kvidrende sang i det tempererede østasiatiske land Japan. Selv om den afrikanske feltgrille Gryllus bimaculatus stammer fra tropiske lande, er den et nyt modeldyr globalt set på grund af dens evne til at regenerere amputerede ben under nymfen og dens udviklingsmåde (kort kimbånd) (Mito og Noji, 2008).
Mange levende organismer i dyreriget er i stand til at genvokse deres kropsdele efter en skade. Eksempler på kropsdele, der kan genvokse, omfatter linsen og halen hos padder, hovedet hos planarier og hjertet hos fisk. Derimod har man længe antaget, at mennesker ikke kan genskabe tabte kropsdele, bortset fra bestemte væv, herunder overhuden, leveren og æggestokkenes overflade efter ægløsning. Derfor er det vigtigt at belyse de molekylære mekanismer, der er involveret i regenereringsprocesser ved hjælp af dyremodeller, der er i stand til at regenerere kropsdele, med henblik på efterfølgende anvendelse i ikke-regenerative menneskelige organer og væv.
I løbet af de sidste 2 år førte sammenlignende genomiske undersøgelser af to planariearter med forskellige regenerative evner til en vellykket regenerering af hoveder ved at reducere beta-catenin-aktiviteten fra ellers ikke-regenerative halefragmenter (Umesono et al., 2013). Undersøgelser af hvirveldyr med evnen til at genoprette lemmer, herunder newts, frøer og salamandre, har vist, at lemmeregenerering sker trinvis. Lemmernes regenereringsproces er opdelt i mindst tre faser: sårheling, dedifferentiering og genopbygning, hvor genopbygningsfasen efterligner den embryonale udvikling (Endo et al., 2004).
Grillebenet består af seks segmenter, der er anbragt langs den proximo-distale (PD) akse: coxa, trochanter, femur, tibia, tarsus og klo (Figur 1). 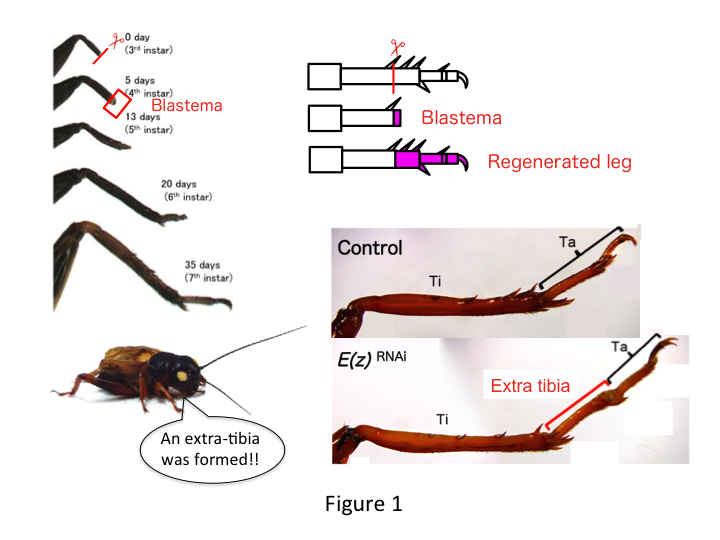 Tarsus er yderligere underopdelt i tre tarsomerer. Når skinnebenet hos nymfen i tredje instar amputeres, regenereres benet og genvinder sin allometriske størrelse og rette form inden det sjette instar (dvs. inden for 20 dage efter amputationen) og får næsten normal voksenstørrelse og -form igen. Snart efter helingen udvikles blastemaet (en pulje af celler, der formerer sig) i den distale region af det amputerede ben. Blastemacellerne formerer sig og danner de manglende strukturer ved hjælp af interkalære processer mellem den mest distale region og den resterende del af benet (French et al., 1976).
Tarsus er yderligere underopdelt i tre tarsomerer. Når skinnebenet hos nymfen i tredje instar amputeres, regenereres benet og genvinder sin allometriske størrelse og rette form inden det sjette instar (dvs. inden for 20 dage efter amputationen) og får næsten normal voksenstørrelse og -form igen. Snart efter helingen udvikles blastemaet (en pulje af celler, der formerer sig) i den distale region af det amputerede ben. Blastemacellerne formerer sig og danner de manglende strukturer ved hjælp af interkalære processer mellem den mest distale region og den resterende del af benet (French et al., 1976).
Tidligere udførte vi en sammenlignende transkriptomanalyse af regenererende og normale amputerede ben af syrener for at profilere mRNA-ekspression, der er forbundet med benregeneration (Bando et al., 2013). Vi fokuserede først på opregulering af Jak/Stat pathway-gener, som er forbundet med immunsystemet. RNA-interferens (RNAi) af generne i denne vej forstyrrede benregenerationen grundigt. I modsætning hertil forårsagede RNAi mod Socs, en undertrykker af cytokinsignalering, benforlængelse. Yderligere eksperimenter viste, at Jak/Stat-vejen fremmer celleproliferation nedstrøms Ds/Fat-vejen.
Derpå undersøgte vi epigenetisk regulering under regeneration af ben fra fårekyllinger. Tetsuya Bando, en seniorforsker i vores gruppe, identificerede et gen for histon H3 lysin 27 (H3K27) methyltransferase, E(z), og et gen for histon H3K27 demethylase, Utx, i G. bimaculatus. Kloning af Gryllus-gener er nu en ukompliceret proces på grund af de oplysninger, der er tilgængelige om grillenes genom (Mito og Noji, personlig kommunikation). Methylering af histon H3K27 af E(z) undertrykker ekspressionen af målgener ved at rekruttere Polycomb-gruppeproteiner. Omvendt fremmer demethylering af det trimethylerede histon H3K27 med Utx genekspressionen. Tetsuya fandt, at transkriptionen af både E(z)- og Utx-generne er opreguleret i blastemacellerne fra amputerede ben (Bando et al., 2013). In situ hybridisering verificerede, at begge gener er ubiquitært transskriberet i de regenererende ben af fårekyllinger, og at begge gener udtrykkes i udviklingsembryoner (Hamada et al., 2015). Immunfarvning på de amputerede små ben efter RNAi af Yoshimasa Hamada (en ph.d.-studerende) bekræftede, at E(z) og Utx bidrager til methylering og demethylering ved henholdsvis histon H3K27me3 under benregeneration.
Derimod fandt Yoshimasa uventet, at det ekstra bensegment dannes efter RNAi mod E(z) (Figur 1). I første omgang var vi ikke i stand til at bestemme identiteten af bensegmentet. Morfologisk set syntes bensegmentet at være en tibia, fordi det havde pigge og sporer, der var karakteristiske for en ægte tibia. Vores indledende hypotese var, at fænotyperne efter RNAi kunne afhænge af amputationsstedet i skinnebenet. Men selv når et ben amputeres i den distale del af lårbenet, fremkommer det ekstra tibia-lignende segment. Mønsterdannelsen langs de antero-posteriore og dorso-ventrale akser forblev uændret, undtagen langs PD-aksen. Vi undersøgte derefter, om amputationsstedet langs PD-aksen i skinnebenet påvirkede den fænotypiske sværhedsgrad. Den ekstra-tibia, der blev dannet, blev længere, jo mere proximalt amputationstederne på tibia var (figur 1). Omvendt resulterede RNAi mod Utx i tab af leddannelse mellem tarsomere 1 (Ta1) og Ta2 (Figur 2) (Figur 2). 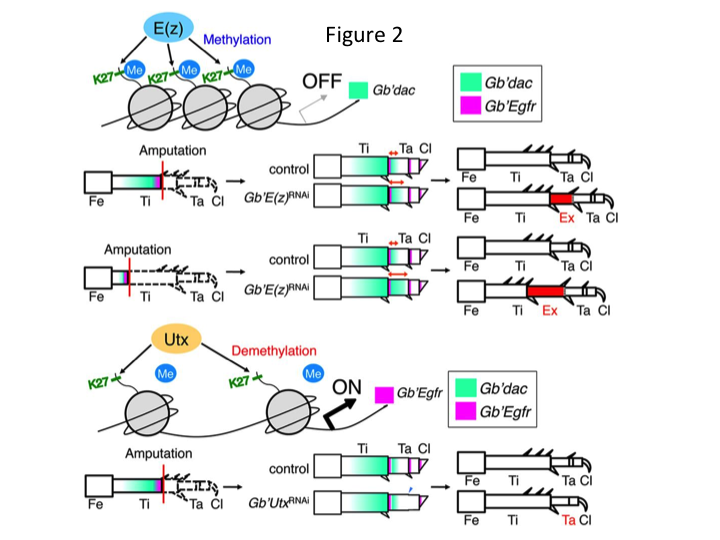 In situ-hybridisering viste, at ekspressionen af benmønstergenerne ændrede sig langs PD-aksen. Specifikt udvidede domænet for dachshund (dac) ekspressionen i E(z)RNAi regenererende ben, mens Egfr-ekspressionen mindskedes i UtxRNAi-ben. Derfor kan E(z) undertrykke dac-ekspression under normal benregeneration, mens Utx inducerer Egfr-ekspression.
In situ-hybridisering viste, at ekspressionen af benmønstergenerne ændrede sig langs PD-aksen. Specifikt udvidede domænet for dachshund (dac) ekspressionen i E(z)RNAi regenererende ben, mens Egfr-ekspressionen mindskedes i UtxRNAi-ben. Derfor kan E(z) undertrykke dac-ekspression under normal benregeneration, mens Utx inducerer Egfr-ekspression.
dac koder for en transkriptionel co-repressor, der er kategoriseret i bengabgener. dac producerer grove positionsværdier langs benets PD-akse og medierer dannelsen af den distale tibia og Ta1 (den proximale tarsomere) under krebsebenregeneration (dac-ekspressionsdomæne er vist med grønt i figur 2) (Ishimaru et al., 2015). Specifikt fremmer dac tibialcelleproliferation. Derfor, fordi RNAi mod E(z) opregulerer dac, kan E(z)-udtryk i blastemacellerne undertrykke den blastemale overproliferation ved at undertrykke ekstra dac-ekspression.
Denne information rejser spørgsmålet om, hvordan E(z) specifikt regulerer dac-ekspression. Endvidere, hvad er den mekanisme, der bestemmer målgenerne for E(z)? E(z) tilhører Polycomb repressive complex 2 (PRC2), som er et af de tre Polycomb group (PcG)-komplekser (Schuettengruber et al., 2007). Under grilleembryogenese undertrykker E(z) den forreste ekspansion af Hox-genekspression og sørger for korrekt identitet i embryoner (Matsuoka et al., 2015). Disse oplysninger indikerer, at målgenerne for E(z) er forskellige afhængigt af den cellulære kontekst. Et DNA-bindingsprotein, Pleiohomeotic (Pho), binder sammen med andre faktorer til Polycomb-responselementerne (PRE) af målgenerne, hvorefter E(z) trimethylerer histon H3K27. Selv om PRE’er kun er blevet identificeret i Drosophila, har meta-analysen af formodede målgener for PcG-proteiner vist, at mange af målgenerne er fælles for fluen, musen og mennesket. dac og Egfr er inkluderet blandt disse gener (Schuettengruber et al., 2007). Således indeholder den regulerende region af kribensis dac-genet sandsynligvis PRE’er, gennem hvilke E(z) epigenetisk regulerer ekspressionen af dac under kribensisbenets regeneration (Figur 2). Den igangværende forskning er fokuseret på at karakterisere funktionerne af Pho-genet og andre PcG-kompleksgener og epigenetiske modifikatorer under Gryllus-benregeneration.
Til sidst, hvorfor forårsager E(z) RNAi ekstra-tibia-dannelse? Et hypotetisk scenarie er, at når skinnebenet amputeres i den proximale position, hvor dac-ekspressionen er lav, tillader Utx-ekspression (som dominerer E(z)-ekspressionen) dac-ekspressionen (figur 3a) at genoprette skinnebenet (figur 3a). 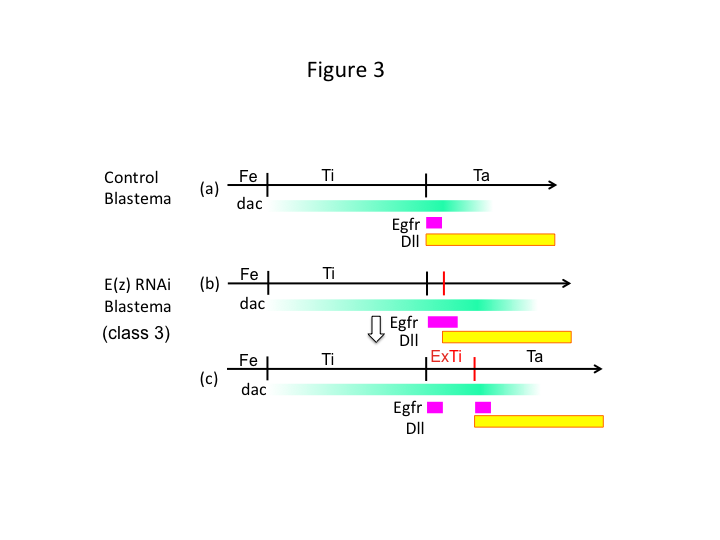 Disse histonmodifikatorer registrerer således positionsværdierne langs amputationsstedets PD-akse og finjusterer ekspressionsniveauet for benmønstergenerne, som f.eks. dac. I tilfælde af E(z) RNAi lige før proximal amputation induceres et intenst dac-ekspression og udvides i det regenererende ben (figur 3b). Distal-less (Dll)-udtrykket, som er et andet benmønstergen, der specificerer det distale domæne af benet (Angelini og Kaufman, 2005), kan forskydes mere distalt afhængigt af det udvidede dac-tryk (Figur 3b). Det Egfr-udtrykkende domæne kan således være opdelt i to dele, hvor (1) Dll-ekspressionen er lav og (2) Dll er høj. Ekstra-tibia dannes sandsynligvis mellem de to forskellige Egfr-udtrykkende domæner ved at interkalere celleproliferation og mønsterdannelse (Figur 3c).
Disse histonmodifikatorer registrerer således positionsværdierne langs amputationsstedets PD-akse og finjusterer ekspressionsniveauet for benmønstergenerne, som f.eks. dac. I tilfælde af E(z) RNAi lige før proximal amputation induceres et intenst dac-ekspression og udvides i det regenererende ben (figur 3b). Distal-less (Dll)-udtrykket, som er et andet benmønstergen, der specificerer det distale domæne af benet (Angelini og Kaufman, 2005), kan forskydes mere distalt afhængigt af det udvidede dac-tryk (Figur 3b). Det Egfr-udtrykkende domæne kan således være opdelt i to dele, hvor (1) Dll-ekspressionen er lav og (2) Dll er høj. Ekstra-tibia dannes sandsynligvis mellem de to forskellige Egfr-udtrykkende domæner ved at interkalere celleproliferation og mønsterdannelse (Figur 3c).
Vores mål er at klarlægge blueprints til “at lave et regenereret ben” ved hjælp af denne attraktive hemimetabolous insektmodel. Disse tegninger forventes at kunne klarlægge, hvordan antallet af bensegmenter bestemmes. Vores slående observationer af RNAi mod E(z), der fører til “ekstra tibia-dannelse”, udgør et vigtigt skridt i retning af at belyse denne proces.
- Mito, T. og Noji, S. (2008). Den toplettede græshoppe Gryllus bimaculatus: En ny model for udviklings- og regenereringsstudier. Cold Spring Harb Protoc, 331-346.
- Umesono, Y., Tasaki, J., Nishimura, Y., Hrouda, M., Kawaguchi, E., Yazawa, S., Nishimura, O., Hosoda, K., Inoue, T. og Agata, K. (2013). Den molekylære logik for planariernes regeneration langs den anterior-posterior akse. Nature 500, 73-76.
- Endo, T., Bryant, S. V. og Gardiner, D. M. (2004). Et trinvis modelsystem til regeneration af lemmer. Dev Biol 270, 135-145.
- French, V., Bryant, P. J., og Bryant, S. V. (1976). Mønsterregulering i epimorfe felter. Science 193, 969-981.
- Bando, T., Ishimaru, Y., Kida, T., Hamada, Y., Matsuoka, Y., Nakamura, T., Ohuchi, H., Noji, S. og Mito, T. (2013). Analyse af RNA-Seq-data afslører involvering af JAK/STAT-signalering under benregeneration hos grillen Gryllus bimaculatus. Development 140, 959-964.
- Hamada, Y., Bando, T., Nakamura, T., Ishimaru, Y., Mito, T., Noji, S., Tomioka, K. og Ohuchi, H. (2015). Benregeneration er epigenetisk reguleret af histon H3K27-methylering i grillen Gryllus bimaculatus. Development 142, 2916-2927.
- Ishimaru, Y., Nakamura, T., Bando, T., Matsuoka, Y., Ohuchi, H., Noji, S. og Mito, T. (2015). Involvering af dachshund og Distal-less i distal mønsterdannelse af krebsebenet under regeneration. Sci Rep 5, 8387.
- Schuettengruber, B., Chourrout, D., Vervoort, M., Leblanc, B. og Cavalli, G. (2007). Genomregulering ved polycomb- og trithoraxproteiner. Cell 128, 735-745.
- Matsuoka, Y., Bando, T., Watanabe, T., Ishimaru, Y., Noji, S., Popadić, A. og Mito, T. (2015). Kortkimede insekter udnytter både den forfødte og afledte måde af Polycomb-gruppe-medieret epigenetisk silencing af Hox-generne. Biol Open 4, 702-709.
- Angelini,R. og Kaufman, T. C. (2005). Insekters vedhæng og sammenlignende ontogenetik. Dev Biol 286, 57-77.
Del
- Emne:*
- Dit navn:*
- Din e-mail:
- Din e-mail:
- *
- Detaljer:*
Indsend rapport
 (1 stemmer)
(1 stemmer)
Tags: Kromatin og epigenetik, syren, regeneration
Kategorier: Kromatin og epigenetik, syren, regeneration
Kategorier: Forskning