În toamnă, greierii prezintă în general cântece de ciripit în țara temperată din Asia de Est, Japonia. Deși greierul de câmp african Gryllus bimaculatus provine din țările tropicale, acesta este un animal model emergent la nivel global datorită capacității sale de regenerare a picioarelor amputate în timpul nimfei și a modului său de dezvoltare (bandă germinală scurtă) (Mito și Noji, 2008).
Multe organisme vii din regnul animal sunt capabile să își regenereze părțile corpului în urma unei leziuni. Printre exemplele de părți ale corpului care pot fi regenerate se numără cristalinul și coada amfibienilor, capul planarilor și inima peștilor. În schimb, s-a presupus mult timp că oamenii nu pot reface părțile corpului pierdute, cu excepția anumitor țesuturi, inclusiv epiderma, ficatul și suprafața ovariană după ovulație. Prin urmare, este important să se elucideze mecanismele moleculare implicate în procesele de regenerare folosind modele animale care sunt capabile să regenereze părți ale corpului pentru aplicarea ulterioară în organele și țesuturile umane care nu se regenerează.
În ultimii 2 ani, studiile genomice comparative a două specii de planari cu abilități regenerative diferite au dus la regenerarea cu succes a capetelor prin reducerea activității beta-cateninei din fragmente de coadă care altfel nu se regenerează (Umesono et al., 2013). Studiile asupra vertebratelor cu capacitatea de a reface membrele, inclusiv tritonii, broaștele și salamandrele, au demonstrat că regenerarea membrelor are loc într-o manieră etapizată. Procesul de regenerare a membrelor este împărțit în cel puțin trei faze: vindecarea rănilor, dediferențierea și redezvoltarea, faza de redezvoltare imitând dezvoltarea embrionară (Endo et al., 2004).
Piciorul de greier este compus din șase segmente care sunt dispuse de-a lungul axei proximo-distale (PD): coxa, trohanterul, femurul, tibia, tarsul și gheara (Figura 1). 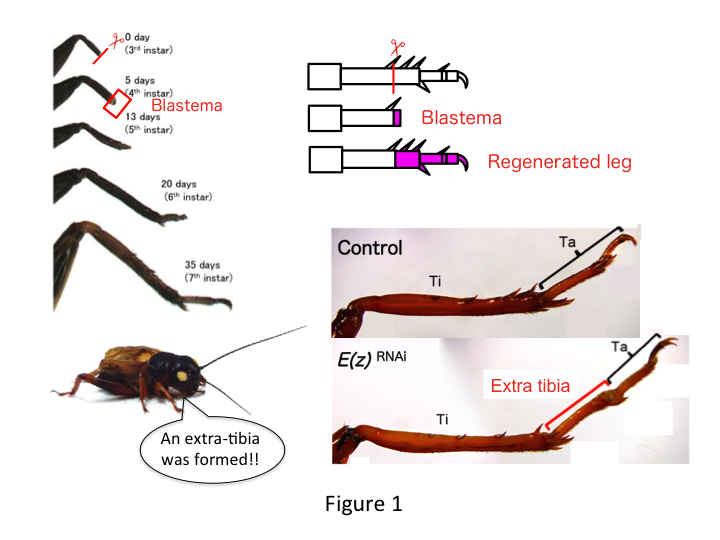 Tarsusul este subdivizat în continuare în trei tarsomeri. Atunci când tibia nimfei de al treilea stadiu este amputată, piciorul se regenerează și își recuperează dimensiunea alometrică și forma corespunzătoare până la al șaselea stadiu (adică în decurs de 20 de zile de la amputare), revenind la dimensiunea și forma aproape normală de adult. La scurt timp după vindecare, blastema (un bazin de celule care proliferează) se dezvoltă în regiunea distală a piciorului amputat. Celulele blastemei proliferează și formează structurile lipsă prin procese intercalare între regiunea cea mai distală și partea rămasă a piciorului (French et al., 1976).
Tarsusul este subdivizat în continuare în trei tarsomeri. Atunci când tibia nimfei de al treilea stadiu este amputată, piciorul se regenerează și își recuperează dimensiunea alometrică și forma corespunzătoare până la al șaselea stadiu (adică în decurs de 20 de zile de la amputare), revenind la dimensiunea și forma aproape normală de adult. La scurt timp după vindecare, blastema (un bazin de celule care proliferează) se dezvoltă în regiunea distală a piciorului amputat. Celulele blastemei proliferează și formează structurile lipsă prin procese intercalare între regiunea cea mai distală și partea rămasă a piciorului (French et al., 1976).
Anterior, am efectuat o analiză comparativă a transcriptomului picioarelor regenerate și a picioarelor amputate normale ale greierilor pentru a profila expresia ARNm asociată cu regenerarea piciorului (Bando et al., 2013). Ne-am axat mai întâi pe reglarea ascendentă a genelor căii Jak/Stat, care sunt legate de sistemul imunitar. Interferența ARN (RNAi) a genelor din această cale a perturbat complet regenerarea picioarelor. În schimb, RNAi împotriva Socs, un supresor al semnalizării citokinelor, a provocat alungirea picioarelor. Experimente suplimentare au arătat că calea Jak/Stat promovează proliferarea celulară în aval de calea Ds/Fat.
Subiectiv, am investigat reglarea epigenetică în timpul regenerării piciorului de greiere. Tetsuya Bando, un cercetător senior din grupul nostru, a identificat o genă pentru metiltransferaza histonei H3 lizina 27 (H3K27), E(z), și o genă pentru demitilaza histonei H3K27, Utx, la G. bimaculatus. Clonarea genelor Gryllus este în prezent un proces simplu datorită faptului că sunt disponibile informații despre genomul greierului (Mito și Noji, comunicare personală). Metilarea histonei H3K27 de către E(z) reprimă expresia genelor țintă prin recrutarea proteinelor din grupul Polycomb. În schimb, demitilarea histonei trimetilate H3K27 de către Utx promovează expresia genelor. Tetsuya a constatat că transcrierea atât a genelor E(z), cât și a genei Utx este suprareglementată în celulele blastematice ale picioarelor amputate (Bando et al., 2013). Hibridizarea in situ a verificat că ambele gene sunt transcrise în mod ubicuu în picioarele regenerate ale greierilor și că ambele gene sunt exprimate în embrionii în curs de dezvoltare (Hamada et al., 2015). Imunocolorarea pe picioarele mici amputate după RNAi de către Yoshimasa Hamada (un doctorand) a confirmat că E(z) și Utx contribuie la metilarea și, respectiv, demetilarea la nivelul histonei H3K27me3, în timpul regenerării picioarelor.
Cu toate acestea, Yoshimasa a constatat în mod neașteptat că segmentul de picior suplimentar se formează după RNAi împotriva lui E(z) (Figura 1). Inițial, nu am reușit să determinăm identitatea segmentului de picior. Din punct de vedere morfologic, segmentul de picior părea a fi o tibie, deoarece avea spini și pinteni care erau caracteristici unei tibii autentice. Ipoteza noastră inițială a fost că fenotipurile după RNAi ar putea depinde de locul amputării tibiei. Cu toate acestea, chiar și atunci când un picior este amputat în partea distală a femurului, apare un segment suplimentar asemănător tibiei. Formarea modelului de-a lungul axelor antero-posterioară și dorso-ventrală a rămas neschimbată, cu excepția axei PD. Am examinat apoi dacă locul amputării de-a lungul axei PD în tibie a influențat severitatea fenotipică. Extra-tibia care s-a format a devenit cu atât mai lungă cu cât locurile de amputare de pe tibie erau mai proximale (figura 1). În schimb, RNAi împotriva Utx a dus la pierderea formării articulației între tarsomerii 1 (Ta1) și Ta2 (Figura 2) (Figura 2). 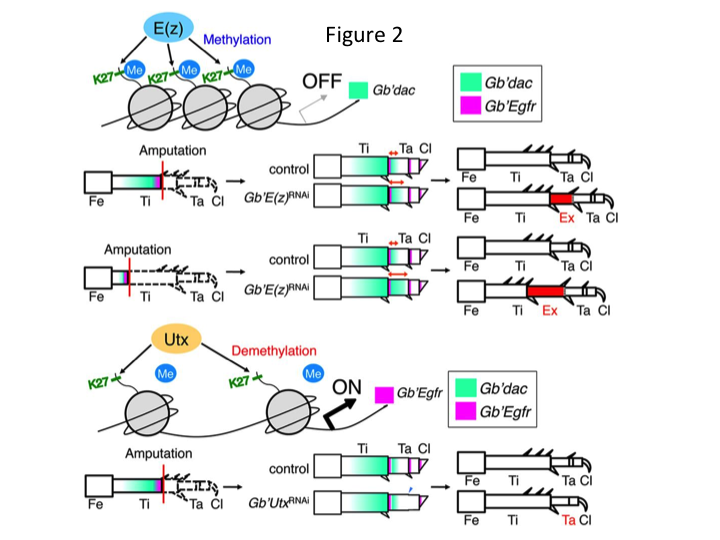 Hibridizarea in situ a arătat că expresia genelor de modelare a piciorului s-a modificat de-a lungul axei PD. Mai exact, domeniul de expresie al dachshund (dac) s-a extins în picioarele în regenerare E(z)RNAi, în timp ce expresia Egfr a scăzut în picioarele UtxRNAi. Prin urmare, E(z) poate reprima expresia dac în timpul regenerării normale a piciorului, în timp ce Utx induce expresia Egfr.
Hibridizarea in situ a arătat că expresia genelor de modelare a piciorului s-a modificat de-a lungul axei PD. Mai exact, domeniul de expresie al dachshund (dac) s-a extins în picioarele în regenerare E(z)RNAi, în timp ce expresia Egfr a scăzut în picioarele UtxRNAi. Prin urmare, E(z) poate reprima expresia dac în timpul regenerării normale a piciorului, în timp ce Utx induce expresia Egfr.
dac codifică un co-represor transcripțional care este categorisit în genele de decalaj ale piciorului. dac produce valori poziționale brute de-a lungul axei PD a piciorului și mediază formarea tibiei distale și a Ta1 (tarsomerul proximal) în timpul regenerării piciorului de greier (domeniul de expresie dac este indicat în verde în figura 2) (Ishimaru et al., 2015). În mod specific, dac promovează proliferarea celulelor tibiale. Prin urmare, deoarece RNAi împotriva lui E(z) stimulează dac, expresia E(z) în celulele blastemei poate suprima supraproliferarea blastemală prin reprimarea expresiei suplimentare a lui dac.
Această informație ridică întrebarea despre modul în care E(z) reglează în mod specific expresia lui dac. Mai mult, care este mecanismul care determină genele țintă ale E(z)? E(z) aparține complexului represiv Polycomb 2 (PRC2), care este unul dintre cele trei complexe din grupul Polycomb (PcG) (Schuettengruber et al., 2007). În timpul embriogenezei greierelor, E(z) reprimă expansiunea anterioară a expresiei genelor Hox și asigură o identitate adecvată în embrioni (Matsuoka et al., 2015). Aceste informații indică faptul că genele țintă ale E(z) diferă în funcție de contextul celular. O proteină de legare a ADN, Pleiohomeotic (Pho), împreună cu alți factori, se leagă de elementele de răspuns Polycomb (PRE) ale genelor țintă, după care E(z) trimetilează histona H3K27. Deși PRE-urile au fost identificate doar la Drosophila, meta-analiza genelor țintă putative pentru proteinele PcG a arătat că multe dintre genele țintă sunt comune la muscă, șoarece și om. dac și Egfr sunt incluse printre aceste gene (Schuettengruber et al., 2007). Astfel, regiunea de reglementare a genei dac a greierului conține probabil PRE-uri, prin care E(z) reglează epigenetic expresia dac în timpul regenerării piciorului de greier (figura 2). Cercetările în curs de desfășurare se concentrează pe caracterizarea funcțiilor genei Pho și a altor gene din complexul PcG și modificatori epigenetici în timpul regenerării piciorului de Gryllus.
În cele din urmă, de ce determină ARNi E(z) formarea de extra-tibia? Un scenariu ipotetic este că, atunci când tibia este amputată în poziția proximală, unde expresia dac este scăzută, expresia Utx (care domină expresia E(z)) permite expresia dac (figura 3a) pentru a reface tibia. 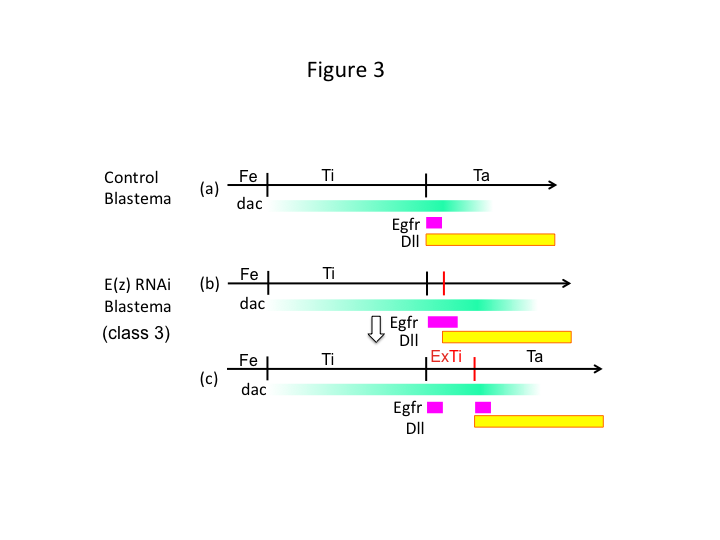 Astfel, acești modificatori histonici detectează valorile poziționale de-a lungul axei PD a locului amputării și reglează fin nivelul de expresie al genelor de modelare a piciorului, precum dac. În cazul RNAi E(z) chiar înainte de amputarea proximală, este indusă o expresie intensă a dac și se extinde în piciorul care se regenerează (figura 3b). Expresia distal-less (Dll), care este o altă genă de distanțare a piciorului care specifică domeniul distal al piciorului (Angelini și Kaufman, 2005), se poate deplasa mai mult distal în funcție de expresia dac extinsă (Figura 3b). Astfel, domeniul care exprimă Egfr poate fi separat în două părți în care (1) expresia Dll este scăzută și (2) Dll este ridicată. Extra-tibia se formează probabil între cele două domenii diferite care exprimă Egfr prin intercalarea proliferării și modelării celulelor (Figura 3c).
Astfel, acești modificatori histonici detectează valorile poziționale de-a lungul axei PD a locului amputării și reglează fin nivelul de expresie al genelor de modelare a piciorului, precum dac. În cazul RNAi E(z) chiar înainte de amputarea proximală, este indusă o expresie intensă a dac și se extinde în piciorul care se regenerează (figura 3b). Expresia distal-less (Dll), care este o altă genă de distanțare a piciorului care specifică domeniul distal al piciorului (Angelini și Kaufman, 2005), se poate deplasa mai mult distal în funcție de expresia dac extinsă (Figura 3b). Astfel, domeniul care exprimă Egfr poate fi separat în două părți în care (1) expresia Dll este scăzută și (2) Dll este ridicată. Extra-tibia se formează probabil între cele două domenii diferite care exprimă Egfr prin intercalarea proliferării și modelării celulelor (Figura 3c).
Obiectivul nostru este de a elucida planurile pentru „fabricarea unui picior regenerat” prin utilizarea acestui model atractiv de insectă hemimetabolatică. Se așteaptă ca planurile să clarifice modul în care este determinat numărul de segmente ale piciorului. Observațiile noastre izbitoare privind RNAi împotriva E(z) care duce la „formarea de tibii suplimentare” reprezintă un pas important spre elucidarea acestui proces.
- Mito, T. și Noji, S. (2008). The Two-Spotted Cricket Gryllus bimaculatus: An emerging Model for Developmental and Regeneration Studies (Un model emergent pentru studii de dezvoltare și regenerare). Cold Spring Harb Protoc, 331-346.
- Umesono, Y., Tasaki, J., Nishimura, Y., Hrouda, M., Kawaguchi, E., Yazawa, S., Nishimura, O., Hosoda, K., Inoue, T. și Agata, K. (2013). Logica moleculară pentru regenerarea planarilor de-a lungul axei antero-posterioare. Nature 500, 73-76.
- Endo, T., Bryant, S. V. și Gardiner, D. M. (2004). Un sistem model etapizat de model pentru regenerarea membrelor. Dev Biol 270, 135-145.
- French, V., Bryant, P. J. și Bryant, S. V. (1976). Reglarea modelelor în câmpurile epimorfe. Science 193, 969-981.
- Bando, T., Ishimaru, Y., Kida, T., Hamada, Y., Matsuoka, Y., Nakamura, T., Ohuchi, H., Noji, S. și Mito, T. (2013). Analiza datelor RNA-Seq dezvăluie implicarea semnalizării JAK/STAT în timpul regenerării picioarelor la greierul Gryllus bimaculatus. Development 140, 959-964.
- Hamada, Y., Bando, T., Nakamura, T., Ishimaru, Y., Mito, T., Noji, S., Tomioka, K. și Ohuchi, H. (2015). Regenerarea picioarelor este reglementată epigenetic de metilarea histonei H3K27 la greierele Gryllus bimaculatus. Development 142, 2916-2927.
- Ishimaru, Y., Nakamura, T., Bando, T., Matsuoka, Y., Ohuchi, H., Noji, S. și Mito, T. (2015). Implicarea Dachshund și Distal-less în formarea modelului distal al piciorului de greier în timpul regenerării. Sci Rep 5, 8387.
- Schuettengruber, B., Chourrout, D., Vervoort, M., Leblanc, B. și Cavalli, G. (2007). Reglarea genomului de către proteinele polycomb și trithorax. Cell 128, 735-745.
- Matsuoka, Y., Bando, T., Watanabe, T., Ishimaru, Y., Noji, S., Popadić, A. și Mito, T. (2015). Insectele cu germeni scurți utilizează atât modul ancestral, cât și cel derivat de reducere la tăcere epigenetică mediată de grupul Polycomb a genelor Hox. Biol Open 4, 702-709.
- Angelini,R. și Kaufman, T. C. (2005). Apendice de insecte și ontogenetica comparativă. Dev Biol 286, 57-77.
Share
- Issue:*
- Your Name:*
- Your Email:*
- Detalii:*
Trimiteți raportul
 (1 voturi)
(1 voturi)
Etichete:
Etichete: Cromatină și epigenetică, greieri, regenerare
Categorii: Cărbunele, regenerare: Cercetare