ADVERTISEMENTS:
I den här artikeln kommer vi att diskutera Platyhelminthes:- 1. Habitat och livsmiljö för Platyhelminthes 2. Platyhelminthes struktur 3. Organ för vidhäftning 4. Kroppsvägg 5. Matsmältningssystemet 6. Utsöndringssystem 7. Andningsorgan 8. Nervsystemet 9. Reproduktionssystemet 10. Utveckling 11. Fylogenetiska överväganden.
- Habitat och livsmiljö för Platyhelminthes:
- Struktur hos Platyhelminthes:
- Häftorgan eller sugorgan hos platyhelminter:
- Kroppsväggen hos Platyhelminthes:
- Förtäringssystemet hos Platyhelminthes:
- Skretoriska systemet hos Platyhelminthes:
- Respiratoriska och cirkulatoriska system hos Platyhelminthes:
- Nervsystemet hos platyhelminter:
- Förökningssystem hos Platyhelminthes:
- Platyhelminternas utveckling:
- Fylogenetiska överväganden om Platyhelminthes:
Habitat och livsmiljö för Platyhelminthes:
Platyhelmintherna är mestadels ekto- eller endoparasitiska och få är frilevande. De frilevande tillhör klassen Turbellaria och lever i sötvatten, dammar, sjöar, vattendrag och källor. En del av dem finns i stränder i tropiska och subtropiska regioner. Trematoda och cestoda är totala parasiter. I vuxenstadiet parasiterar de på ryggradsdjur och i larvstadiet förekommer de som parasiter på ryggradslösa djur.
Struktur hos Platyhelminthes:
ADVERTISEMENTS:
Fylums medlemmar är vanligtvis avlånga till utseendet. Polykladerna är breda bladliknande medan bandmaskarna är platta och bandliknande. Kroppens kontur är i allmänhet enkel men vissa trematoder erbjuder bisarra konturer. Bandmaskarnas kroppar består av ett antal fyrkantiga eller rektangulära segment som kallas proglottider.
Den främre proglottiden är minst och den bakersta proglottiden är störst. Det innebär att proglottidens storlek ökar i anteroposterior riktning. Förekomsten av proglottider ger bandmaskarna ett segmenterat tillstånd. Ett slags pseudometamerism förekommer hos Procerodes lobata där vissa av de inre organen upprepas.
Förre och bakre ändar samt dorsala och ventrala ytor är lätt igenkännliga. Ofta avgränsas den främre änden från resten av kroppen genom förekomsten av ett ”huvud” följt av en inskränkt ”hals”.
I vissa former saknas definitivt huvud men den främre delen kan upptäckas av sinnesorganen eller genom att dess rörelse är riktad framåt under förflyttning. Ventralytan bär munnen och genitalöppningarna, när de finns.
RÅDGÅNGAR:
Storleken på platyhelminter varierar från mikroskopiska till extremt långsträckta former som kan vara 10-15 meter långa (bandmaskar). De flesta av medlemmarna har små till måttliga dimensioner. Platyhelminter är i allmänhet färglösa eller vita till färgen, fritt levande former är vita, bruna, gråa eller svarta till färgen. Vissa polyklader och landplanarier har ljusa färger som är arrangerade i mönster.
Häftorgan eller sugorgan hos platyhelminter:
Platyhelmintherna har en mängd olika organ för vidhäftning och fastsättning. Acetabulum eller sugorgan i form av ”sug” är mycket vanligt hos vuxna plattmaskar. Hos fläckar finns det två sugkoppar på den ventrala sidan av kroppen. En av dessa sugkoppar är belägen på den främre sidan av kroppen och dess position är mer eller mindre fast.
Men den andra som kallas bakre suggapparaten är inte konstant i sin position. Hos Paramphistomum är suggapparaten längst bak i position, hos Echinostoma och Fasciola är positionen förskjuten mer framåt.
I bandmaskar finns de adhesiva organen i form av rännor eller koppar och är belägna i den cefala änden. Ofta finns det krokar i närheten av dessa spår eller i den evigt buckala kammaren för att hjälpa till med förankringen. Sugare förekommer också hos vissa frilevande Planaria.
Kroppsväggen hos Platyhelminthes:
Platyhelminthes saknar exo- eller endoskelett och kroppen är därför mjuk. Epidermis är enkelskiktad och i vissa fall är den syncytial. Epidermis kan vara cilierad helt eller delvis. Det subepidermala lagret består av muskulatur av cirkulära, longitudinella och sneda fibrer. Muskelfibrerna är släta. Alla utrymmen mellan organen är fyllda med packningsceller, som kallas parenkym.
De hårda delarna hos Platyhelminthes är krokarna och taggarna. Många tidigare arbetare ansåg att kroppen hos monogeneaner, digeneaner och cestoder är täckt av ”kutikula”.
Non-ciliation hos dessa former ger det falska intrycket av kutikula under ljusmikroskopi. Men elektronmikroskopiska studier har visat att kroppens yttre skikt är epidermis. Cellerna i detta skikt innehåller mikrokondrier och förblir kontinuerliga med de underliggande cellerna.
Förtäringssystemet hos Platyhelminthes:
Förmningsorganen saknas hos turbellarorden Acoela och hos cestoderna. Hos Turbellaria och Trematoda representeras matsmältningssystemet av mun, svalg och tarm som slutar blint och som sådant liknar hela dispositionen av matsmältningsorganen i hög grad den hos antozoer och ctenophorer.
Munnen är i primitivt tillstånd belägen ungefär i mitten av ventrallinjen, men hos många plattmaskar är positionen förskjuten framåt längs mitten av ventrallinjen.
ADVERTIVERINGAR:
Munnen saknas hos endoparasitiska rhabdocoeler. Farynx är stomodaeal till sin natur och är ett kraftigt muskulärt rör. Den uppvisar variation inom fylumet. Tarmen uppvisar en stor variation i form. Den kan vara en enkel säck eller ha komplicerade förgreningar och underförgreningar. Analöppningar förekommer sällan.
Skretoriska systemet hos Platyhelminthes:
Platyhelminthernas utsöndringssystem består huvudsakligen av längsgående vattenkärl eller längsgående utsöndringskanaler som har ett antal specialiserade utsöndringsceller, kallade flamceller eller flamlökar, som tillsammans kallas protonephridier. Vattenkärlens placering i kroppen erbjuder en stor variation.
I triklerna finns det två längsgående kärl (kanaler) som öppnar sig utåt genom många porer (fig. 14.28G). Rhabdocoels har två laterala eller ett enda mediankärl (fig. 14.28E). Mediankärlet öppnar sig utåt genom en enda por som ligger i kroppens bakre ände.
I former där det finns två kärl finns öppningarna antingen på den ventrala sidan eller i svalget. Hos vissa former förenas de två kärlen i det bakre området och öppnar sig utåt genom en enda medianöppning. Den huvudsakliga nephridialkanalen är kraftigt förgrenad och det finns accessoriska utsöndringsporer förutom nephridioporerna (fig. 14.28F).
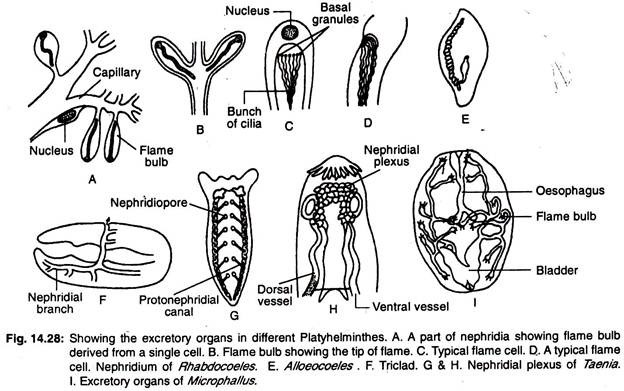
I monogena trematoder finns det två längsgående utsöndringskanaler och de två kanalerna öppnas separat på dorsalytan. Kanalerna utvidgas till att bilda utsöndringssäckar vid sina ändar.
I andra trematoder är utsöndringskanalerna baktill förbundna med varandra genom en tvärgående kanal och öppnas utåt genom en enda medianöppning. Den tvärgående kanalen dialaterar och bildar i vissa fall en utsöndringsblåsa. De två längsgående kanalerna kan förenas med varandra i det bakre området och kan ha en enda öppning.
I cestoder finns det fyra längsgående utsöndringsvestiler. Utsöndringskärlen är förbundna med ett tvärgående kärl som ligger i scolexet för att bilda nephridinal plexus (fig. 14.28H) och mynnar i en kontraktila utsöndringsblåsor som ligger i den sista proglottiden.
Numera protonephridier och ett stort antal nephridioporer finns hos sötvattensturbeller eftersom inflödet av överflödigt vatten förhindras i kroppen.
Respiratoriska och cirkulatoriska system hos Platyhelminthes:
Åtgärder:
Dessa två system saknas helt i fylumet. Hos vissa trematoder har förekomsten av ett system av rör, kallat lymfsystemet, beskrivits. Rören anses ha osäkra funktioner.
Nervsystemet hos platyhelminter:
Det huvudsakliga nervcentrumet eller hjärnan ligger i huvudet som ett par hjärnganglier. Från hjärnan utgår flera ganglifierade längssnören. Av dessa longitudinella snören blir ett par mest iögonfallande och resten blir obetydliga. Många tvärgående förbindelser uppstår mellan de longitudinella banden och hela nervsystemet får ett steglöst utseende.
Sensoriska organ som ocelli eller ögon finns hos turbellarior och monogena trematoder. De är talrika hos polyklader och två till fyra hos Rhabdocoels och monogena trematoder. Ögonen finns antingen som pigmentceller eller i form av koppar som innehåller pigmentceller och sensoriska celler. Chemo- och tangoreceptorer är allmänt förekommande och statocystor förekommer hos Acoela och några få andra former.
Förökningssystem hos Platyhelminthes:
Med undantag av ett fåtal Turbellaria och Trematoda är platyhelminter hermafroditer. De manliga och kvinnliga reproduktionsorganen i varje individ är separata och öppnas utvändigt genom egna porer eller genom en gemensam genitalöppning. I vissa fall mynnar gonodukterna ut i matsmältningskanalen och könscellerna frigörs genom munnen.
ÅTERKÄNNANDE:
I vissa plattmaskar finns en extra honlig por eller vaginalporer som tjänar under kopulationen. Gonoporerna är vanligtvis ventralt placerade men är ibland dorsalt placerade. Hos bandmaskar är gonoporerna lateralt placerade.
Manliga fortplantningsorgan består av testiklar som i primitivt tillstånd är många och utspridda. Antalet testiklar är reducerat till en eller två hos många och hos Acoela saknas definitiva gonader och kanaler. Vasa efferentia, när de finns, motsvarar antalet testiklar. I allmänhet finns det ett par vasa deferentia som förenas och mynnar ut i den komplicerade kopulationsapparaten.
Kopulationsapparaten består av en ständigt vridbar cirrus eller en utskjutande penis som är beväpnad med taggar eller krokben. Olika körtlar är knutna till den. Vesiklar i enkelt eller parat tillstånd är ofta närvarande och fungerar som reservoar för lagring av spermier. De kvinnliga könsorganen består av en eller två äggstockar. När äggledarna är parade smälter de ihop distalt och bildar en gemensam äggledare som mynnar ut i kopulationsapparaten.
Kopulationsapparaten består av en säck, sädesblåsan eller kallad sädesbursa eller kopulationsbursa. Olika körtlar som hjälper till att bilda äggskal och producera sekretoriska ämnen är också knutna till kopulationsapparaten. En lång tubulär eller förgrenad livmoder för att samla mogna ägg utgör en iögonfallande del av fortplantningssystemet.
Den kvinnliga gonaden hos platyhelminter är märklig eftersom den är tydligt differentierad i två zoner, den egentliga äggstocken och äggula- eller vitellinkörtlarna.
Gulan ingår aldrig i äggets struktur som hos andra djur, utan produceras som abortiva ägg som ingår i äggkapseln eller skalet för att ge mat åt det embryo som utvecklas. Hos cestoderna upprepas fortplantningsorganen i varje proglottid. Hos unga proglottider är organen rudimentära medan de är högt utvecklade hos gravida proglottider.
Platyhelminternas utveckling:
REDSKAP:
Och även om de är hermafroditer är det vanligt med korsbefruktning. Befruktningen sker internt och befruktade, skalade ägg som innehåller embryon fälls utåt. Acoela och Polyclads saknar gulkörtlar och utvecklingen är direkt. Hos Polyclads produceras en frisimmande larv som antas förebåda Trochophore.
Larven kallas Mullers larv eller Protrochula (fig. 14.29). Den är oval till formen och bär åtta framträdande armar som är besatta med långa cilier som bildar ett sammanhängande band. Den allmänna kroppsytan är täckt av små cilier.
Munkavsnittet är beläget i mitten av den ventrala linjen och tre ögon finns i den främre delen av dorsalytan. Under utvecklingen absorberas de cilierade armarna. Hos andra grupper är embryoutvecklingen kraftigt modifierad och komplicerad. 
Fylogenetiska överväganden om Platyhelminthes:
Det finns ingen enhällig uppfattning bland zoologer när det gäller plattmaskarnas ursprung. Det finns flera åsikter om plattmaskarnas ursprung.
1. Lang (1881) föreslog ctenophore- polyclad-teorin. Enligt denna teori kan polykladdade turbellarior ha utvecklats från ctenophorer genom Platyctenea. Mullers larv av polyklad har vissa likheter med ctenophorer.
De likartade dragen är:
ADVERTISERINGAR:
(i) Likheter i symmetrierna mellan ctenophorerna och larven.
(ii) 8 ektodermala ciliala kammar hos Mullers larv kan jämföras med de meridionala kammarplattorna hos ctenophorerna.
(iii) Statocystan hos turbellarior kan jämföras med ctenophorerna.
Denna teori gynnar inte mycket stöd bland zoologer.
2. Planula larva-teorin säger att turbellarior har vissa likheter med planula larverna hos nässeldjur. Så många zoologer som Hyman (1951), Jagersten (1955) och Hadzi (1963) anser att plattmaskar har utvecklats från ett planula-liknande förfädersystem och att Acoela är den mest primitiva gruppen bland platyhelminter (fig. 14.27A).
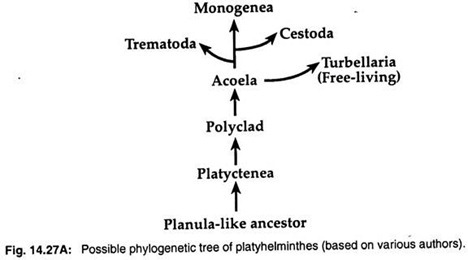
Molekylära data och kladistiska analyser har föreslagit ett monofyletiskt ursprung för de tre parasitgrupperna. De tre parasitära klasserna Trematoda, Monogenea och Cestoda har utvecklats från fritt levande turbellarior.
3. Återigen säger enterocoel-teorin att alla bilaterala djur i grunden är coelomerade, och acoelomerade plattmaskar har utvecklats från coelomerade förfäder genom förlust av håligheten sekundärt. Denna teori är inte godtagbar för zoologerna eftersom den är svår att förklara vissa aspekter, t.ex. förändringen från coelomat till acoelomat tillstånd och från bilateral till radiell symmetri.