Lärandemål
- Identifiera historiska och potentiella orsaker till höga utrotningsfrekvenser
Antalet arter på planeten, eller i något geografiskt område, är resultatet av en jämvikt mellan två evolutionära processer som ständigt pågår: artbildning och utrotning. Båda är naturliga ”födelse-” och ”dödsprocesser” i makroevolutionen. När artrikedomen börjar bli större än utdöendet kommer antalet arter att öka. På samma sätt kommer antalet arter att minska när utdöendet börjar bli större än artrikedomen. Under hela jordens historia har dessa två processer fluktuerat – ibland har de lett till dramatiska förändringar i antalet arter på jorden, vilket återspeglas i fossilregistret (figur 1).
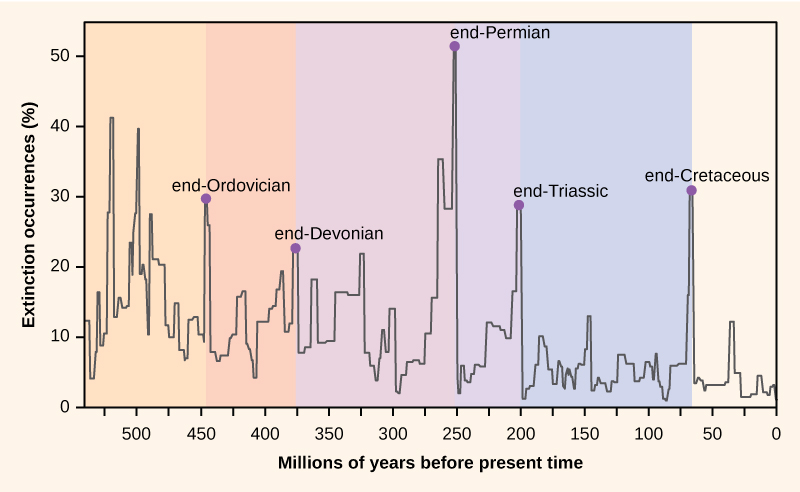
Figur 1. Procentuella utdöenden som återspeglas i fossilregistret har fluktuerat under hela jordens historia. Plötsliga och dramatiska förluster av biologisk mångfald, så kallade massutdöenden, har inträffat fem gånger.
Paleontologer har identifierat fem strata i fossilregistret som tycks uppvisa plötsliga och dramatiska (mer än hälften av alla existerande arter försvinner från fossilregistret) förluster i biologisk mångfald. Dessa kallas massutdöenden. Det finns många mindre, men ändå dramatiska, utdöenden, men de fem massutdöendena har lockat till sig mest forskning. Man kan hävda att de fem massutdöendena bara är de fem mest extrema händelserna i en kontinuerlig serie av stora utdöenden under hela Phanerozoikum (sedan 542 miljoner år tillbaka). I de flesta fall är de antagna orsakerna fortfarande kontroversiella; den senaste händelsen verkar dock vara tydlig.
Registrerade massutdöenden
Fossilregistret över massutdöendena låg till grund för att definiera perioder av den geologiska historien, så de inträffar vanligen vid övergångspunkten mellan geologiska perioder. Övergången i fossil från en period till en annan återspeglar den dramatiska förlusten av arter och det gradvisa uppkomsten av nya arter. Dessa övergångar kan ses i bergsskikten. Tabell 1 innehåller uppgifter om de fem massutdöendena.
| Tabell 1. Massutdöenden | ||
|---|---|---|
| Geologisk period | Namn på massutdöendet | Tid (miljoner år sedan) |
| Ordovician-Silurian | slut.Ordovicium O-S | 450-440 |
| Senaste devonier | slut-Devonian | 375-360 |
| Permian-Triassic | end-Permian | 251 |
| Triassic-Jurassic | end-Trias | 205 |
| Kristid-Paleogen | end-Kristid K-Pg (K-T) | 65.5 |
Det ordovicisk-siluriska utdöendet är det första registrerade massutdöendet och det näst största. Under denna period dog cirka 85 procent av de marina arterna (få arter levde utanför haven) ut. Huvudhypotesen för dess orsak är en period av istid och därefter uppvärmning. Utdöendet består i själva verket av två utdöende händelser som är åtskilda med cirka 1 miljon år. Den första händelsen orsakades av nedkylning och den andra händelsen berodde på den efterföljande uppvärmningen. Klimatförändringarna påverkade temperaturer och havsnivåer. Vissa forskare har föreslagit att en gammastrålning, orsakad av en närliggande supernova, var en möjlig orsak till utdöendet mellan ordovicium och silur. Gammastrålningen skulle ha avlägsnat jordens skyddande ozonskikt och gjort det möjligt för intensiv ultraviolett strålning från solen att nå jordens yta, vilket kan förklara de klimatförändringar som observerades vid den tidpunkten. Hypotesen är mycket spekulativ, och utomjordiska influenser på jordens historia är en aktiv forskningslinje. Återhämtningen av den biologiska mångfalden efter massutdöendet tog mellan 5 och 20 miljoner år, beroende på plats.
Det sena devoniska utdöendet kan ha skett under en relativt lång tidsperiod. Det verkar ha påverkat främst marina arter och inte så mycket växter eller djur som bebor landmiljöer. Orsakerna till detta utdöende är dåligt kända.
Det sena permiska utdöendet var det största i livets historia. Man skulle faktiskt kunna hävda att jorden blev nästan helt utan liv under detta utdöende. Uppskattningar visar att 96 procent av alla marina arter och 70 procent av alla landlevande arter försvann. Det var vid den här tiden som till exempel trilobiterna, en grupp som överlevde utdöendet i Ordovicium-Silurien, dog ut. Orsakerna till detta massutdöende är oklara, men den främsta misstänkta orsaken är en långvarig och utbredd vulkanisk aktivitet som ledde till en skenande global uppvärmning. Haven blev till stor del anoxiska och det marina livet kvävdes. Det tog 30 miljoner år innan mångfalden av landlevande tetrapoder återhämtade sig efter utdöendet i slutet av Perm. Det permiska utdöendet förändrade dramatiskt jordens sammansättning av biologisk mångfald och evolutionens förlopp.
Orsakerna till det triassisk-jurassiska utdöendet är inte klara, och forskarna argumenterar för hypoteser som inkluderar klimatförändringar, asteroidnedslag och vulkanutbrott. Utdöendet inträffade strax innan superkontinenten Pangaea splittrades, även om nyare forskning tyder på att utdöendena kan ha inträffat mer gradvis under hela Trias.
Orsakerna till utdöendet i slutet av kritaperioden är de som är bäst förstådda. Det var under detta utdöende för cirka 65 miljoner år sedan som majoriteten av dinosaurierna, den dominerande ryggradsdjursgruppen under miljontals år, försvann från planeten (med undantag för en theropod-klad som gav upphov till fåglar).
Orsaken till detta utdöende förstås nu som ett resultat av en kataklysmisk nedslag av en stor meteorit, eller asteroid, utanför kusten av vad som nu är Yucatánhalvön. Denna hypotes, som först föreslogs 1980, var en radikal förklaring som byggde på en kraftig ökning av nivåerna av iridium (som kommer in i vår atmosfär från meteorer i en ganska konstant takt men som i övrigt saknas på jordens yta) i det stenlager som markerar gränsen mellan krit- och paleogenperioden (figur 2). Denna gräns markerade dinosauriernas försvinnande i fossil samt många andra taxa. Forskarna som upptäckte iridiumspiken tolkade den som ett snabbt inflöde av iridium från rymden till atmosfären (i form av en stor asteroid) snarare än en avtagande avlagring av sediment under den perioden. Det var en radikal förklaring, men rapporten om en nedslagskrater av lämplig ålder och storlek 1991 gjorde hypotesen mer trovärdig. Nu stöder ett överflöd av geologiska bevis teorin. Återhämtningstiderna för den biologiska mångfalden efter utdöendet i slutet av krita är kortare, i geologisk tid, än för utdöendet i slutet av perm, i storleksordningen 10 miljoner år.
En annan möjlighet, som kanske sammanfaller med nedslaget av asteroiden från Yucatan, var omfattande vulkanism som började bildas för cirka 66 miljoner år sedan, ungefär samtidigt som nedslaget av asteroiden från Yucatan, i slutet av kritan. Lavaströmmarna täckte över 50 procent av det som nu är Indien. Utsläppet av vulkaniska gaser, särskilt svaveldioxid, under bildandet av fallen bidrog till klimatförändringen, som kan ha framkallat massutdöendet.
Praktikfråga

Figur 2. Iridiumband (kredit: USGS)
År 1980 upptäckte Luis och Walter Alvarez, Frank Asaro och Helen Michels, över hela världen, en topp i koncentrationen av iridium i det sedimentära lagret vid K-Pg-gränsen. Dessa forskare antog att denna iridiumspik orsakades av ett asteroidnedslag som resulterade i K-Pg massutdöendet. I figur 2 är iridiumskiktet det ljusa bandet.
Vetenskapsmännen mätte den relativa förekomsten av ormbunkssporerna ovanför och under K-Pg-gränsen i detta stenprov. Vilket av följande påståenden representerar troligen deras resultat?
- En mängd ormbunkssporerna från flera arter hittades under K-Pg-gränsen, men ingen hittades ovanför.
- En mängd ormbunkssporerna från flera arter hittades ovanför K-Pg-gränsen, men ingen hittades nedanför.
- En rikedom av ormbunkssporerna hittades både över och under K-Pg-gränsen, men endast en art hittades under gränsen, och många arter hittades ovanför gränsen.
- Många arter av ormbunkssporerna hittades både ovanför och under gränsen, men det totala antalet sporer var större nedanför gränsen.
Det pleistocena utdöendet
Det pleistocena utdöendet är ett av de mindre utdöendena och ett nyligen inträffat utdöende. Det är välkänt att den nordamerikanska, och i viss mån eurasiska, megafaunan – stora ryggradsdjur – försvann mot slutet av den senaste istiden. Utdöendet tycks ha skett under en relativt begränsad tidsperiod för 10 000-12 000 år sedan. I Nordamerika var förlusterna ganska dramatiska och omfattade bl.a. den ulliga mammuten (med en kvarvarande population som existerade fram till för cirka 4 000 år sedan i isolering på Wrangel Island i Kanada), mastodonten, jättebävern, jättelika markandsloddar, sabel-tandade katter och den nordamerikanska kamelen, för att nämna några. I början av 1900-talet föreslog forskare för första gången möjligheten att överdriven jakt orsakade den snabba utrotningen av dessa stora djur. Forskningen om denna hypotes fortsätter idag.
I allmänhet korrelerade tidpunkten för de pleistocena utdöendena med ankomsten av paleomänniskor, kanske redan för 40 000 år sedan, och inte med klimatförändringar, vilket är den viktigaste konkurrerande hypotesen för dessa utdöenden. Utdöendet började i Australien för cirka 40 000-50 000 år sedan, strax efter det att människorna anlände till området: ett pungdjurslöv, en jättelik wombat på ett ton och flera jättelika känguruarter försvann. I Nordamerika dog nästan alla stora däggdjur ut för 10 000-12 000 år sedan. Allt som finns kvar är de mindre däggdjuren som björnar, älgar, älgar och pumor. På många avlägsna öar i havet dog många arter ut i samband med människans ankomst. Alla öar hade inte stora djur, men när det fanns stora djur tvingades de ofta att dö ut. Madagaskar koloniserades för cirka 2 000 år sedan och de stora däggdjur som levde där dog ut. Eurasien och Afrika uppvisar inte det här mönstret, men de upplevde inte heller någon nytillkommen ankomst av jägar- och samlarmänniskor. Människorna anlände snarare till Eurasien för hundratusentals år sedan. Detta ämne är fortfarande ett område för aktiv forskning och hypoteser. Det verkar klart att även om klimatet spelade en roll var det i de flesta fall människans jakt som påskyndade utrotningarna.
Högtidliga utrotningar
Den sjätte massutrotningen, eller den holocena massutrotningen, tycks ha börjat tidigare än vad man tidigare trott och beror till stor del på de störande aktiviteterna av den moderna Homo sapiens. Sedan början av den holocena perioden finns det många nyligen inträffade utdöenden av enskilda arter som finns registrerade i mänskliga skrifter. De flesta av dessa sammanfaller med de europeiska koloniernas expansion sedan 1500-talet.
Ett av de tidigare och allmänt kända exemplen är dronfågeln. Den udda duvliknande fågeln levde i skogarna på Mauritius (en ö i Indiska oceanen) och utrotades omkring 1662. Dodo jagades för sitt kött av sjömän och var ett lätt byte eftersom den närmade sig människor utan rädsla (dronten hade inte utvecklats tillsammans med människor). Grisar, råttor och hundar som de europeiska fartygen förde med sig till ön dödade också drontens ungar och ägg.
Steller’s sea cow dog ut 1768; den var besläktad med manaten och levde troligen en gång i tiden längs Nordamerikas nordvästkust. Stellers sjöko upptäcktes först av européer 1741 och överjagades för kött och olja. Den sista sjökoan dödades 1768. Det blir alltså bara 27 år mellan sjökossans första kontakt med européer och artens utdöende!
Sedan 1900 har en rad olika arter dött ut, bland annat följande:
- År 1914 dog den sista levande passagerarduvan i ett zoo i Cincinnati, Ohio. Denna art hade en gång i tiden mörkat himlen i Nordamerika under sina flyttningar, men den var överjagad och drabbades av förlust av livsmiljöer till följd av att skogar röjdes för jordbruksmark.
- Carolinasparakiten, som en gång i tiden var vanlig i östra USA, dog ut 1918. Den drabbades av habitatförlust och jagades för att förhindra att den åt frukt från fruktträdgårdar. (Parakiten åt frukt från fruktträdgårdar eftersom dess inhemska föda förstördes för att ge plats åt jordbruksmark.)
- Det japanska sjölejonet, som bebodde ett stort område runt Japan och Koreas kust, utrotades på 1950-talet på grund av fiskare.
- Den karibiska munksälen var spridd över hela Karibiska havet men drevs till utrotning genom jakt 1952.
Detta är bara några av de registrerade utrotningarna under de senaste 500 åren. Internationella naturvårdsunionen (IUCN) upprätthåller en förteckning över utdöda och hotade arter som kallas rödlistan. Listan är inte fullständig, men den beskriver 380 utdöda arter av ryggradsdjur efter 1500 e.Kr. varav 86 av dem drevs ut av överjakt eller överfiske.
Uppskattningar av utrotningsfrekvenser i nutid
Uppskattningar av utrotningsfrekvenser försvåras av att de flesta utrotningar troligen sker utan att man observerar dem. Det är troligt att människor märker att en fågel eller ett däggdjur har dött ut, särskilt om de har jagats eller använts på något annat sätt. Men det finns många organismer som är av mindre intresse för människor (inte nödvändigtvis av mindre värde) och många som är obeskrivna.
Bakgrundsutdöendet uppskattas till cirka en per miljon arter per år (E/MSY). Om man till exempel antar att det finns cirka tio miljoner arter, förväntas tio arter dö ut varje år (varje år motsvarar tio miljoner arter per år).
En samtida uppskattning av utrotningsfrekvensen använder sig av de utdöenden som finns i den skriftliga dokumentationen sedan år 1500. Bara för fåglar ger denna metod en uppskattning på 26 E/MSY. Detta värde kan dock vara en underskattning av tre skäl. För det första skulle många arter inte ha beskrivits förrän långt senare under tidsperioden, så deras förlust skulle ha gått obemärkt förbi. För det andra ökar antalet nyligen utdöda ryggradsdjursarter eftersom utdöda arter nu beskrivs från skelettrester. Och för det tredje är vissa arter troligen redan utdöda även om naturvårdare är ovilliga att benämna dem som sådana. Om man tar hänsyn till dessa faktorer ökar den uppskattade utrotningsfrekvensen till närmare 100 E/MSY. Den beräknade utrotningsfrekvensen i slutet av århundradet är 1 500 E/MSY.

Figur 3. Studier har visat att antalet förekommande arter ökar med habitatets storlek. (kredit: modifiering av Adam B. Smiths arbete)
En andra metod för att uppskatta utrotningsfrekvensen i nutid är att korrelera förlust av arter med förlust av livsmiljöer genom att mäta förlust av skogsareal och förstå förhållandet mellan arter och areal. Förhållandet mellan art och areal är den hastighet med vilken nya arter ses när den undersökta arealen ökar. Studier har visat att antalet förekommande arter ökar när öns storlek ökar. Detta fenomen har också visat sig gälla i andra öliknande livsmiljöer, t.ex. bergstopparna i Venezuela, som är omgivna av tropisk skog. Om man vänder på detta förhållande och minskar livsmiljöns areal, kommer också antalet arter som lever där att minska. Uppskattningar av utrotningsfrekvenser baserade på förlust av livsmiljöer och förhållandet mellan arter och arealer har visat att om 90 procent av livsmiljöerna försvinner, förväntas 50 procent av arterna dö ut. Uppskattningar av arters areal har lett till beräkningar av arters utrotningsfrekvens på omkring 1 000 E/MSY och högre. I allmänhet visar de faktiska observationerna inte på en sådan förlust, och det har föreslagits att utrotningen fördröjs. Nyligen utförda arbeten har också ifrågasatt tillämpligheten av förhållandet mellan art och areal när man uppskattar förlusten av arter. I detta arbete hävdas att förhållandet mellan art och areal leder till en överskattning av utrotningsfrekvensen. Ett bättre förhållande att använda kan vara förhållandet mellan endemiska arter och områden. Genom att använda denna metod skulle uppskattningarna sjunka till omkring 500 E/MSY under det kommande århundradet. Observera att detta värde fortfarande är 500 gånger högre än bakgrundsvärdet.
Att prova det
Bidraga!
Förbättra den här sidanLär dig mer