Immunsystemet
Immunsystemet består av vävnader, celler och molekyler vars primära funktion är att upptäcka, reagera på och eliminera patogener och omvandlade celler.
Innate and Adaptive Immune Response
Det finns två huvudkomponenter i immunsystemet – 1) det medfödda immunsystemet och 2) det adaptiva immunsystemet. Det medfödda immunsystemet fungerar som den första försvarslinjen och upptäcker patogener via könscellerkodade mönsterigenkänningsreceptorer, bildar inget minne och reagerar snabbt (inom minuter till timmar). Det adaptiva immunsystemet reagerar långsamt (över flera dagar), använder antigenspecifika receptorer som genomgår flera genomflyttningar under utvecklingen och bildar immunologiskt minne – det ”lär sig” av det första angreppet och förbereder kroppen för att bekämpa framtida exponering för liknande skadligt material. Båda typerna av immunsvar kan antingen drivas av makromolekyler i den extracellulära vätskan eller av aktivering av specifika immunceller. Dessa reaktioner kallas humoral immunitet respektive cellmedierad immunitet.
Under paraplyet cellmedierat immunsvar stratifierar immuncellerna oftast utifrån om svaret är medfött eller adaptivt. Det medfödda immunförsvaret består huvudsakligen av fagocyterande celler, såsom neutrofiler, makrofager, naturliga mördarceller, basofiler och andra som riktar in sig på farliga invaderande organismer. Cellerna i det adaptiva immunförsvaret är T-celler och B-celler. Natural killer (NK) T-celler har egenskaper från både medfödda och adaptiva immunceller. Dessutom kan de flesta andra celltyper, som inte är specifika för immunsystemet, hysa inneboende medfödda immunfunktioner i form av cytoplasmatiska receptorer och signal- och effektormolekyler, t.ex. medlemmar av RIG-I-, STING- och NLR-familjerna. Som vi kommer att se vidare på uppstår celler i båda systemen från olika myeloida och lymfoida linjer av hematopoetiska stamceller.
Integrerade komponenter i immunsystemet
Mogna immunceller differentieras från det som kallas hematopoetiska stamceller. Dessa är pluripotenta odifferentierade progenitorceller som finns i benmärg, perifert blod och placenta. Dessa hematopoetiska stamceller kan differentiera till den vanliga myeloida progenitorcellen eller den vanliga lymfoida progenitorcellen.
Lymfoida celler
Gemensamma lymfoida progenitorceller kan differentieras till fyra huvudpopulationer av lymfocyter som kan särskiljas genom uttryck av receptorer på cellytan: T-celler, B-celler, naturliga mördarceller (NK) och NK-T-celler.
T-celler
T-celler spelar en viktig roll i den cellmedierade immuniteten. Dessa celler identifieras genom uttrycket av CD3 T-cellers signaleringskedjor.
T-celler uppstår från lymfoida progenitorceller som ursprungligen utvecklas från hematopoetiska stamceller i benmärgen. När den lymfoida progenitorn bestämmer sig för att bli en T-cell migrerar den från benmärgen till thymus (därav namnet T-cell). Thymus ger den rätta mikromiljön där T-cellerna kan utvecklas till sina olika subtyper, som definieras av de specifika receptorer som uttrycks på cellytan. Mognande T-celler i de tidiga stadierna, eller tymocyter, saknar receptorer för både CD4 och CD8 och kallas därför dubbelnegativa (DN) celler. DN-cellerna genomgår sedan genetiska omarrangemang och mutationer i gener som kodar för alfa- och beta- T-cellsreceptorerna (TCR), vilket leder till att de börjar följa en linjeförbindelse där slutresultatet är en T-cellsreceptor med specifik affinitet för ett unikt antigen.
Thymocyterna selekteras sedan, både positivt och negativt, för att säkerställa att de känner igen ”främmande” antigener men inte värdantigen, som presenteras som peptider via MHC på cellytan. Endast celler som klarar båda testerna triggas sedan att uttrycka CD4 och CD8 och blir dubbelpositiva celler. I slutet av mognadsprocessen kommer T-cellerna antingen att uttrycka CD4 eller CD8 (och förlora uttrycket av det andra) och därmed bli enkelpositiva celler. Dessa mogna CD4- eller CD8-positiva T-celler släpps sedan ut i blodomloppet och befinner sig till en början i naivt tillstånd, vilket innebär att de ännu inte har bundit sig till en viss T-cellsubtyp.
Om den mognande tymocyten visar en intermediär mängd affinitet för MHC-peptider från det egna könet under mognaden kan den väljas ut för att bli en regulatorisk T-cell (Treg) på grund av sin förmåga att känna igen skadliga ”egna” antigener. T-celler i blodet eller i perifera vävnader kan också lokalt induceras till att bli Treg-celler. Tregcellernas viktigaste uppgift är att upprätthålla tolerans mot självantigener samt att begränsa T-effektorcellers funktion och proliferation.
När de väl anländer till blodet eller den perifera vävnaden kan naiva T-celler induceras till att bli T-hjälparceller. Antigenpresenterande celler (APC), t.ex. makrofager, dendritiska celler och B-celler, kommer att visa antigenet för T-cellen, och om dess receptor har affinitet kommer T-cellen att känna igen antigenet. Typiskt sett är CD4+ T-celler lämpade för att känna igen peptidantigener som är bundna till MHC-proteiner av klass II. När peptid-MHC-komplexet väl känns igen kommer T-cellen att aktivera en rad interna vägar som gör det möjligt att verifiera antigenkännandet. Först då kommer T-hjälparcellen att föröka sig och utöka poolen av tillgängliga celler som är specifika för det skadliga antigenet, t.ex. olika bakterier och parasiter.
När T-hjälparcellerna har känt igen sitt antigen och aktiverats kommer de att frisätta en mängd cytokiner, som är molekyler som signalerar till andra immunceller hur de på lämpligt sätt ska reagera på den skadliga molekylen. Det finns flera undergrupper av hjälpande T-celler, bland annat Th1, Th2 och Th17. Dessa undergrupper producerar och utsöndrar olika cytokiner som hjälper till att skräddarsy immunsvaret beroende på typen av patogen. Th1-celler förlitar sig till exempel på effektorcytokinet IFN-γ för att signalera till makrofagerna att smälta och förstöra det skadliga materialet, medan Th2-cellerna utsöndrar en kombination av IL-4, IL-5, IL-9, IL-10 och IL-13 för att trigga basofiler, eosinofiler och andra celler att angripa infektionen. Den specifika mängden och sorterna av cytokiner avgör det lämpliga svaret på infektionen eller den skadliga cellen. T-celler kan också reagera på celler från det egna jaget genom att bli cytotoxiska T-celler (eller naturliga mördar-T-celler). Typiskt sett är dessa CD8+-celler som känner igen virusinfekterade celler eller tumörceller. Naturliga mördar-T-celler (NKT) kommer att diskuteras närmare i ett senare avsnitt. Dessa och andra typer av T-celler utför sina olika uppgifter eller effektorfunktioner på olika sätt för att känna igen skadligt material och rekrytera andra immunceller för att förstöra det, samt för att bygga upp ett varaktigt immunsvar mot samma typ av angrepp.
T-cellreceptorns interaktiva signalväg granskas i detalj här.
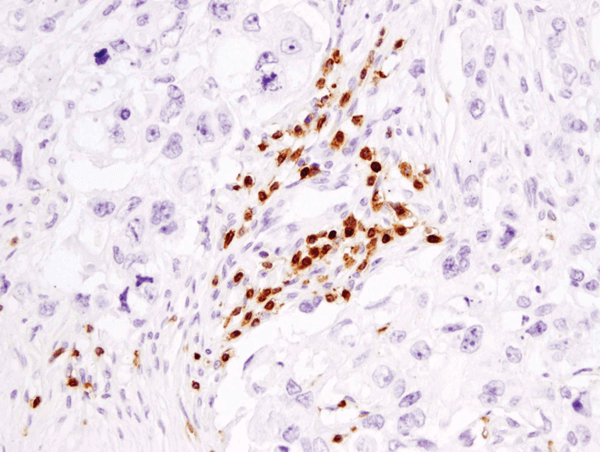
CD3ε (D7A6E™) XP® Rabbit mAb #85061: IHC-analys av paraffinininbäddat humant bröstcancer med #85061.
CD3ε (D7A6E™) XP® Rabbit mAb #85061: IHC-analys av paraffinembeddat humant bröstcancer med hjälp av #85061.
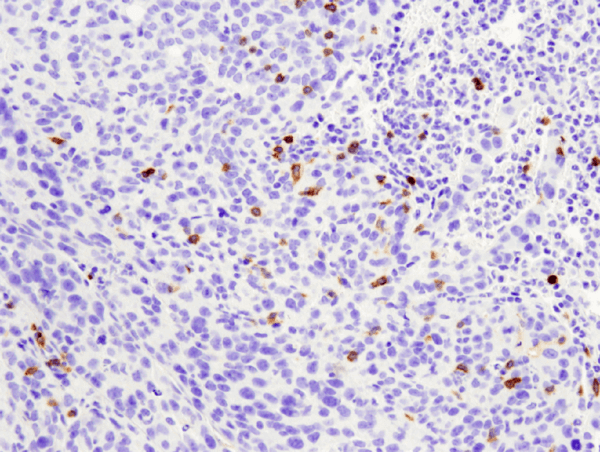
CD4 (D7D2Z) Kanin mAb #25229: IHC-analys av paraffinininbäddad 4T1 metastaserande tumör i musens lunga med hjälp av #25229.
CD4 (D7D2Z) Kanin mAb #25229: IHC-analys av paraffinininbäddad 4T1 metastaserande tumör i muslunga med hjälp av #25229.
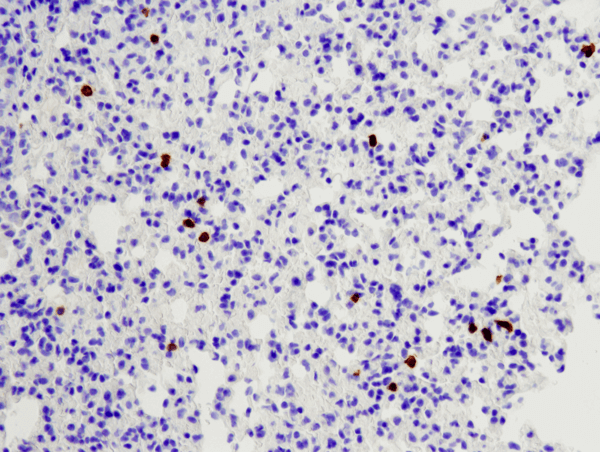
CD8α (D4W2Z) XP® Rabbit mAb (Mouse Specific) #98941: IHC-analys av paraffininbäddad muslunga med hjälp av #98941.
CD8α (D4W2Z) XP® Rabbit mAb (Mouse Specific) #98941: IHC-analys av paraffininbäddad muslunga med hjälp av #98941.
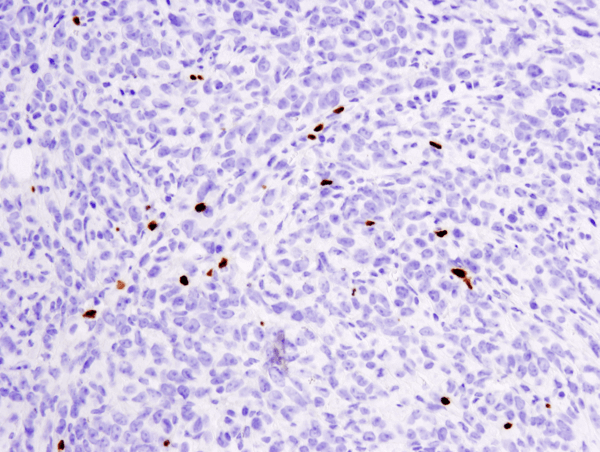
FoxP3 (D6O8R) Rabbit mAb #12653: IHC-analys av paraffinininbäddad 4T1 metastatisk tumör i muslunga med hjälp av #12653.
FoxP3 (D6O8R) Rabbit mAb #12653: IHC-analys av paraffinininbäddad 4T1-metastaserande tumör i muslunga med hjälp av #12653.
B-celler
B-celler är de kritiska cellerna i blodet som förmedlar produktionen av antikroppar. När skadligt material finns i blodet känns det igen som ett antigen av specifika receptorer på B-cellerna. Efter att ha bearbetat antigenet och med hjälp av T-celler mognar sedan B-cellen till plasmaceller som utsöndrar antikroppar. B-celler kan också spela en roll i presentationen av antigener och cytokinutsöndring. Både B-cellernas utveckling och olika funktioner kommer att diskuteras.
15 % av de leukocyter som finns i perifert blod är B-celler. B-celler producerar immunoglobuliner (Ig), som är antigenbindande proteiner (även kallade antikroppar) bestående av två identiska tunga kedjor och två identiska lätta kedjor. Dessa immunoglobuliner innehåller specifika antigenbindningsställen som varierar i aminosyrasekvens från en antikroppsmolekyl till en annan. Dessa platser gör att varje antikropp är specifik för att känna igen ett visst antigen.
Som nämnts ovan differentieras B-celler från hematopoetiska celler som finns i benmärgen. Det är i benmärgen som yt-Ig-receptorer (antikroppar) byggs upp genom en process av genomfördelning. B-celler använder sig av somatisk sammanfogning av olika gener på en mängd kromosomer som kodar för de olika delarna av de tunga och lätta kedjorna som utgör Ig. B-cellerna drivs mot differentiering genom produktion av IL-7 av benmärgens stromaceller. Dessa B-celler under utveckling följer ett program av sekventiella omarrangemang av tunga och lätta kedjor som förmedlas av olika receptoruttryck på cellytan. Uttrycket av en specifik pre-B-cellreceptor förhindrar till exempel vissa genomflyttningar på orelaterade kromosomer för att säkerställa att den mogna B-cellen kan uttrycka en viss Ig-ytreceptor som specifikt kan känna igen ett enskilt antigen. Denna process kallas allelisk uteslutning. Felaktig sammansättning av Ig-tunga kedjor och lätta kedjor kan framkalla sekundära omarrangemang för receptorredigering.
Förutom cellytans uttryck av en Ig-receptor innehåller B-celler också transmembranproteiner som kan inducera intracellulära signaler efter att ett antigen binder till den extracellulära delen av Ig-receptorn. B-celler använder liknande intracellulära signalkaskader som T-celler som inducerar transkriptionella förändringar som leder till proliferation och mognad. I synnerhet kan Ig-receptorns intracellulära domäner fosforyleras av src-familjen av kinaser, vilket initierar en kaskad som vidarebefordras av tyrosinkinaset Syk och ett länkprotein som kallas BLINK till fosfolipas C och guaninutbytesfaktorer, för att slutligen aktivera proteinkinas C, mobilisera kalcium och aktivera MAP-kinas på ett Ras/Rac-beroende sätt.
En unik egenskap hos B-celler är deras förmåga att genomgå isotypbyte. Detta avser specifikt en process av DNA-omläggning som kan generera något olika tunga kedjor. Dessutom kan alternativ splicing av exoner för tunga kedjor göra det möjligt för Ig att förbli bundet till cellytan eller att utsöndras om den transmembranära exonen splicas ut. Alternativ splicing och isotypbyte induceras av cytokiner från T-celler. Dessutom kan T-celler inducera somatiska mutationer som leder till förändringar i de antigenbindande platserna i B-cellernas Ig. En självkorrigerande B-cellsfunktion kan upptäcka om dessa till synes slumpmässiga mutationer leder till en förlust av affinitet för antigenet och initierar ett program för celldöd. Antigener som aktiverar både T-celler och B-celler utlöser således T-cellsmedierad B-cellsmognad genom isotypbyte och somatiska mutationer, vilka båda är starkt förknippade med B-cellsminnet.
Minnessvar kännetecknas av en snabb och omfattande Ig-produktion med hög affinitet mot sådana antigener. Utvecklingen av dessa minnessvar är avgörande för att vaccinering ska lyckas. De kan dock också öka autoimmunitet och allergiska reaktioner.
T-cellsoberoende B-cellsaktivering av ett antigen kan också äga rum; denna typ av aktivering är dock kopplad till ett svagt immunminne. Dessutom måste dessa antigener vara polymera för att kunna aktivera B-celler.
NB: Cytokiner, eller interleukiner (IL), är utsöndrade molekyler som verkar på transmembranreceptorer på cellytan. Vid bindning aktiverar dessa cytokiner intracellulära signaltransduktionsvägar som vanligtvis är beroende av Jak-STAT-signalering.
B-cellreceptorns interaktiva signalväg granskas i detalj här.
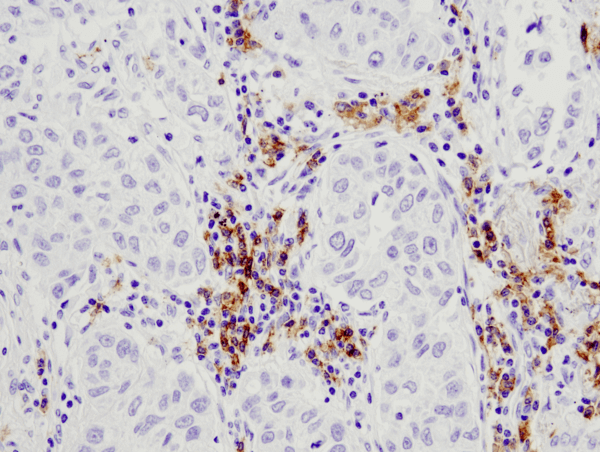
CD19 (D4V4B) XP® Rabbit mAb #90176: IHC-analys av paraffinininbäddat humant lungkarcinom med #90176.
CD19 (D4V4B) XP® Rabbit mAb #90176: IHC-analys av paraffininbäddat humant lungkarcinom med hjälp av #90176.
NK-celler
Naturliga mördarceller, eller NK-celler, är en annan celltyp som härrör från den lymfoida linjen. I likhet med B-celler utvecklas de i benmärgen på ett cytokin- och benmärgsstromalcellsberoende sätt. De utgör en liten andel av det perifera blodet och har inga antigenspecifika receptorer. I stället förlitar sig NK-cellerna på en komplex uppsättning transmembranreceptorer. De innehåller särskilt hämmande cellytereceptorer som känner igen HLA-molekyler av klass I och hämmas därför av egna MHC-molekyler och dödar endast celler som har nedreglerat klass I HLA-uttryck. Därför tjänar de det viktiga syftet att rikta in sig på virusinfekterade celler och tumörceller vars klass I HLA-uttryck är nedreglerat för att undvika att dödas av CD8+ natural killer T-celler (NK-T-celler). NK-celler kan också aktiveras på ett antikropps-, interferon- eller cytokinberoende sätt och kan därför spela en stor roll i kampen mot tumörer. NK-cellernas verkningsmekanism bygger på att små granuler som innehåller granzymer och perforin frigörs från deras cytoplasma. När dessa proteiner frigörs kan de skapa porer och bryta ner intracellulära proteiner för att inducera apoptos. NK-celler sägs därför vara cytotoxiska.
NK-T-celler är en speciell undertyp av T-celler som uttrycker vissa receptorer och markörer som vanligtvis uttrycks av NK-celler i kombination med en begränsad repertoar av T-cellsreceptorer. Därför delar denna unika subtyp egenskaper med både T-celler och NK-celler. Till skillnad från NK-celler, som utvecklas i märgen, utvecklas NK-T-celler i thymus.
Myeloida celler
Gemensamma myeloida progenitorceller, som är ättlingar till hematopoetiska stamceller, ger upphov till flera olika typer av myeloida celler, inklusive makrofager, dendritiska celler och flera andra medfödda immunceller och blodceller.
Makrofager
Makrofager kan utlösas för att känna igen antigener, t.ex. skadade celler eller främmande material, för destruktion på begäran. Makrofager finns i de flesta vävnader och reagerar vid behov på infektioner och döende celler. Det erkända materialet förstörs via fagocytos i makrofagen, vilket ger cellerna deras namn (”storätare” på grekiska). Makrofager antar olika former när de finns på olika platser och kan utföra ytterligare funktioner utöver fagocytos.
Vid vävnadsskada eller patogeninfektion rekryteras monocyter i blodet till den drabbade vävnaden och differentieras till makrofager. Beroende på vävnadslokaliseringen finns det olika typer av makrofager, t.ex. Kupffer-celler i levern, alveolära makrofager i lungan, mikroglia i hjärnan osv. Dessa olika typer av makrofager kommer alla från monocyter men specialiserar sin funktion till den inhemska vävnaden. Det mesta av den allmänna fagocytosfunktionen utförs av makrofager i den bofasta vävnaden. Förutom att fagocytera döda celler och främmande material kan makrofager även signalera till andra immunceller via cytokiner. I viss utsträckning utför makrofagerna den kritiska funktionen antigenpresentation och arbetar följaktligen tillsammans med T-celler för att stödja den adaptiva immuniteten. Dessutom kan makrofager utsöndra cytokiner som IL-12 och spela en roll i lokala immunsvar, medan andra utsöndrar stora mängder IL-10, som förmedlar deras roll i vävnadsreparation. De ”stora ätarna” spelar alltså en mängd olika roller i immunförsvaret utöver sin huvuduppgift, nämligen fagocytos.
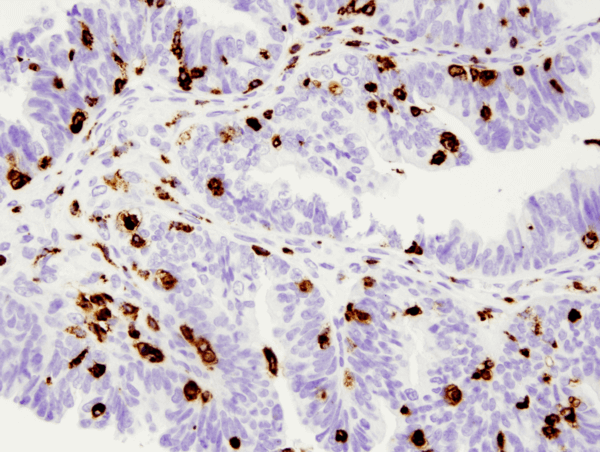
CD68 (D4B9C) XP® Rabbit mAb #76437: IHC-analys av paraffinininbäddat humant seröst papillärt karcinom i äggstocken med #76437.
CD68 (D4B9C) XP® Rabbit mAb #76437: IHC-analys av paraffininbäddat humant seröst papillärt karcinom i ovariet med #76437.
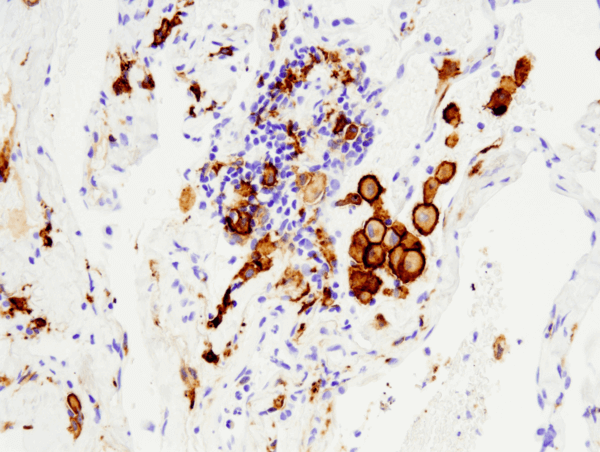
CD163 (D6U1J) Kanin-MAb #93498: IHC-analys av paraffinininbäddat humant icke-småcellig lungcancer med hjälp av #93498, utförd på Leica® Bond™ Rx.
CD163 (D6U1J) Kanin mAb #93498: IHC-analys av paraffinininbäddat humant icke-småcellig lungcancer med hjälp av #93498 utförd på Leica® Bond™ Rx.
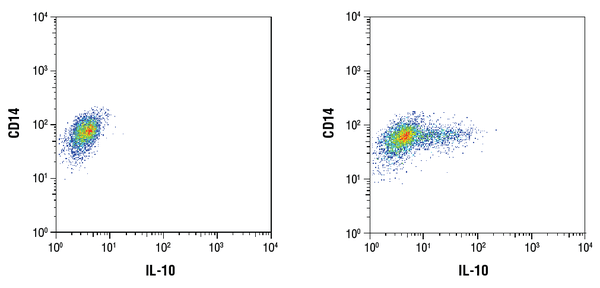
IL-10 (D13A11) XP® Rabbit mAb #12163: Flödescytometrisk analys av renade CD14+ mononukleära celler från mänskligt perifert blod obehandlade (vänster) eller behandlade med LPS (100ng/mL, 16 timmar; höger) med hjälp av en CD14-antikropp och #12163. Brefeldin A #9972 (300ng/mL) tillsattes till obehandlade och behandlade celler efter 1 timmes LPS-stimulering. Anti-rabbit IgG (H+L), F(ab’)2 Fragment (Alexa Fluor® 647 Conjugate) #4414 användes som sekundär antikropp.
IL-10 (D13A11) XP® Rabbit mAb #12163: Flödescytometrisk analys av renade CD14+ mononukleära celler från mänskligt perifert blod obehandlade (vänster) eller behandlade med LPS (100ng/mL, 16 timmar; höger) med hjälp av en CD14-antikropp och #12163. Brefeldin A #9972 (300ng/mL) tillsattes till obehandlade och behandlade celler efter 1 timmes LPS-stimulering. Anti-rabbit IgG (H+L), F(ab’)2 Fragment (Alexa Fluor® 647 Conjugate) #4414 användes som sekundär antikropp.
Dendritiska celler
I likhet med makrofager har dendritiska celler förmågan att uppsluka cellulärt och främmande material, men i stället för att smälta det för att avlägsna det bearbetar de dendritiska cellerna materialet och presenterar det som antigener för T-celler. Som sådana fungerar dendritiska celler som antigenpresenterande celler, eller accessoriska celler, och förmedlar därmed information om patogener mellan det medfödda och det adaptiva immunsystemet. Dendritiska celler har fått sitt namn från sina tydliga cellprojektioner som sträcker sig ut och tar tag i skadligt material för att smälta det.
De omogna dendritiska cellerna, som uppkommer från hematopoietiska progenitorer i benmärgen, genomgår till en början en ”inlärningsfas” där de undersöker sitt närområde för att hitta främmande material och initierar ett igenkänningssvar via mönsterigenkänningsreceptorer (en vanlig uppsättning av dessa receptorer är de toll-liknande receptorerna). Dessa omogna dendritiska celler kan finnas på olika platser med tillgång till den yttre miljön, t.ex. i näsan, magen, lungorna och till och med i huden. När antigenet känns igen vandrar de dendritiska cellerna till lymfkörteln där de tränar T-cellerna i att känna igen antigenet och reagera därefter. Dendritiska celler är de enda antigenpresenterande cellerna som aktiverar både minnes- och naiva T-celler. Toll-like-receptorer är en välstuderad klass av receptorer som finns på makrofager och dendritiska celler och som förmedlar igenkänning och reaktion på främmande patogener. Förutom att interagera direkt med T-celler kan dendritiska celler även signalera via cytokiner som IL-12, som verkar för att förbereda immunförsvaret för den efterföljande attacken mot det skadliga materialet. Andra typer av dendritiska celler kan utsöndra cytokiner som signalerar att de ska rekrytera fler makrofager för att rensa upp en överbelastning av avfallsmaterial. Som de viktigaste cellmediatorerna för antigenpresentation spelar dendritiska celler således en oumbärlig roll i samspelet mellan det medfödda och det adaptiva immunsystemet.
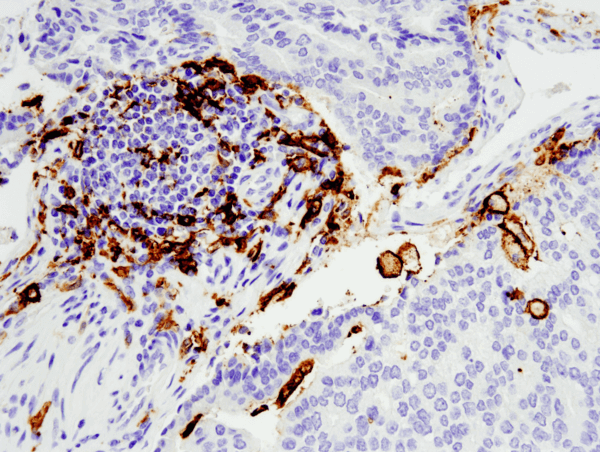
CD11c (D3V1E) XP® Rabbit mAb #45581: IHC-analys av paraffininbäddat humant prostatacancer med hjälp av #455812 utförd på Leica® Bond™ Rx.
CD11c (D3V1E) XP® Rabbit mAb #45581: IHC-analys av paraffininbäddat humant prostatacancer med hjälp av #455812 utförd på Leica® Bond™ Rx.
Andra celler i det medfödda immunförsvaret och signalering
Andra celler i det medfödda immunförsvaret inkluderar mastceller, som fungerar vid sårläkning och bidrar till att ge ett skydd mot patogener. När de utlöses av en patogen kan mastcellerna utsöndra cytokiner samt granuler som innehåller histamin och heparin, vilka medierar ytterligare immunreaktioner såsom rekrytering av makrofager och utvidgning av blodkärl. Neutrofiler, eosinofiler och basofiler kallas tillsammans för granulocyter på grund av de karakteristiska granulerna i deras cytoplasma. I neutrofiler verkar granulerna för att frigöra giftiga föreningar som riktar sig mot patogener som svampar och bakterier.
Immunologi i hälsa och sjukdom
Cancerimmunologi
Att utnyttja immunceller för att bekämpa cancer har blivit ett mycket hett forskningsämne på grund av framgångar i kliniken. En ny inriktning för den nuvarande immunterapin är att rikta in sig på signaleringskontrollproteiner, t.ex. PD-1, som normalt förhindrar aktivering och respons från T-cellerna. Adoptiv cellterapi är ett annat spännande tillvägagångssätt som innebär immunoeditering av T-celler som hämtas från patienten. T-cellerna är konstruerade för att uttrycka en specifik chimär antigenreceptor som är specifik för patientens cancerceller. T-cellerna återinfunderas sedan i patienten och är nu programmerade att söka upp cancercellerna och döda dem, i enlighet med den normala T-cellsfunktionen. Dessa och andra metoder för att manipulera immunsystemet så att det angriper patientens skadliga cancerceller utgör en banbrytande klass av behandlingar för att behandla cancer.