ADVERTISEMENTS:
En este artículo hablaremos de los Platyhelminthes:- 1. Hábitat y Habitat de los Platyhelminthes 2. Estructura de los Platyhelminthes Estructura de los platelmintos 3. Órganos de Adhesión 4. Pared del cuerpo 5. Sistema digestivo 6. Sistema excretor 7. Sistema respiratorio 8. Sistema nervioso 9. Sistema reproductor 10. Desarrollo 11. Consideraciones filogenéticas.
- Hábitat y forma de vida de los platelmintos:
- Estructura de los Platyhelminthes:
- Órganos de Adhesión o Chupadores en Platyhelminthes:
- Pared del cuerpo de Platyhelminthes:
- Sistema digestivo de los platelmintos:
- Sistema excretor de los platelmintos:
- Sistemas respiratorio y circulatorio de los platelmintos:
- Sistema nervioso de los platelmintos:
- Sistema reproductivo de Platyhelminthes:
- Desarrollo de los platelmintos:
- Consideraciones filogenéticas de los platelmintos:
Hábitat y forma de vida de los platelmintos:
Los platihelmintos son en su mayoría ecto o endoparásitos y pocos son de vida libre. Los de vida libre pertenecen a la clase Turbellaria y viven en agua dulce, estanques, lagos, arroyos y manantiales. Algunos de ellos se encuentran en las costas de las regiones tropicales y subtropicales. Los Trematoda y los Cestoda son parásitos totales. En estado adulto parasitan a los vertebrados y en estado larvario se presentan como parásitos de los animales invertebrados.
Estructura de los Platyhelminthes:
ADVERTENCIAS:
Los miembros del filo suelen tener un aspecto alargado. Los policlados tienen forma de hoja ancha, mientras que las tenias son planas y con forma de cinta. El contorno del cuerpo es simple en general, pero algunos trematodos ofrecen un contorno extraño. El cuerpo de las tenias está formado por una serie de segmentos cuadrangulares o rectangulares llamados proglótidos.
El proglótido anterior es el más pequeño y el posterior el más grande. Es decir, el tamaño de la proglótida aumenta en la dirección anteroposterior. La presencia de proglótidos confiere a las tenias una condición segmentada. En Procerodes lobata se encuentra una especie de pseudometamerismo en el que se repiten algunos de los órganos internos.
Los extremos anterior y posterior así como las superficies dorsal y ventral son fácilmente reconocibles. A menudo el extremo anterior se distingue del resto del cuerpo por la presencia de una «cabeza» seguida de un «cuello» constreñido.
En algunas formas carecen de cabeza definida pero el extremo anterior puede detectarse por los órganos sensoriales o por su movimiento dirigido hacia delante durante la locomoción. La superficie ventral lleva la boca y las aberturas genitales, cuando están presentes.
ADVERTENCIAS:
Los tamaños de los platihelmintos varían desde formas microscópicas hasta formas extremadamente alargadas de hasta 10-15 metros de longitud (Tenias). La mayoría de los miembros son de dimensiones pequeñas a moderadas. Los platihelmintos en general son incoloros o blancos, las formas de vida libre son de color blanco, marrón, gris o negro. Algunos policlados y planarios terrestres son de colores brillantes dispuestos en patrones.
Órganos de Adhesión o Chupadores en Platyhelminthes:
Los platihelmintos poseen una gran variedad de órganos de adhesión y de sujeción. El acetábulo u órgano succionador en forma de «ventosa» es muy común en los gusanos planos adultos. En los gusanos planos, hay dos ventosas en la parte ventral del cuerpo. Una de estas ventosas está situada en la parte anterior del cuerpo y su posición es más o menos fija.
Mientras que la otra llamada ventosa posterior no es constante en su posición. En Paramphistomum la ventosa es de posición más posterior, en Echinostoma y Fasciola la posición está desplazada más anteriormente.
En las tenias los órganos adhesivos están presentes en forma de surcos o copas y están situados en el extremo cefálico. A menudo hay ganchos cerca de estos surcos o en la cámara bucal eversible para ayudar al anclaje. Las ventosas también se dan en algunas Planaria de vida libre.
Pared del cuerpo de Platyhelminthes:
Los Platyhelminthes carecen de exo o endoesqueleto y como tal el cuerpo es blando. La epidermis es de una sola capa y en algunos casos es sincitial. La epidermis puede ser ciliada en su totalidad o en parte. La capa subepidérmica está formada por una musculatura de fibras circulares, longitudinales y oblicuas. Las fibras musculares son lisas. Todos los espacios entre los órganos están rellenos de células de relleno, llamadas parénquima.
Las partes duras en los Platyhelminthes son los ganchos y las espinas. Muchos trabajadores anteriores consideraron que el cuerpo de los monogéneos, digéneos y céstodos está cubierto por «cutícula».
La falta de conciliación en estas formas da la falsa impresión de que hay cutícula bajo la microscopía óptica. Pero los estudios de microscopía electrónica han revelado que la capa externa del cuerpo es la epidermis. Las células de esta capa contienen microcondrias y permanecen continuas con las células subyacentes.
Sistema digestivo de los platelmintos:
El sistema digestivo está ausente en el orden de los turbelarios Acoela y en los cestodos. En Turbellaria y Trematoda el sistema alimentario está representado por la boca, la faringe y el intestino, que termina de forma ciega, por lo que la disposición completa de los órganos digestivos se asemeja mucho a la de los antozoos y ctenóforos.
La boca en estado primitivo está situada aproximadamente en la mitad de la línea ventral, pero en muchos platelmintos la posición se desplaza anteriormente a lo largo de la línea ventral media.
ADVERTENCIAS:
La boca está ausente en los rabdóceros endoparásitos. La faringe es de naturaleza estomodal y es un tubo muscular fuerte. Muestra variación en el filo. El intestino muestra una amplia variación en su forma. Puede ser un saco simple o puede tener ramificaciones y sub-ramificaciones complicadas. Las aberturas anales rara vez están presentes.
Sistema excretor de los platelmintos:
El sistema excretor de los platihelmintos consiste principalmente en vasos de agua longitudinales o canales excretores longitudinales que tienen un número de células excretoras especializadas, llamadas células de llama o bulbos de llama, denominadas colectivamente protonefridios. La disposición de los vasos de agua en el interior del cuerpo ofrece una amplia gama de variaciones.
En los triclados hay dos vasos longitudinales (canales) que se abren al exterior por numerosos poros (Fig. 14.28G). Los rabdoceles tienen dos vasos laterales o un solo vaso mediano (Fig. 14.28E). El vaso mediano se abre al exterior por un único poro situado en el extremo posterior del cuerpo.
En las formas en las que hay dos vasos, las aberturas están en la parte ventral o en la faringe. En algunas formas los dos vasos se unen en la región posterior y se abren al exterior por una única abertura mediana. El canal nefridial principal está muy ramificado y hay poros excretores accesorios además del nefridioporo (Fig. 14.28F).
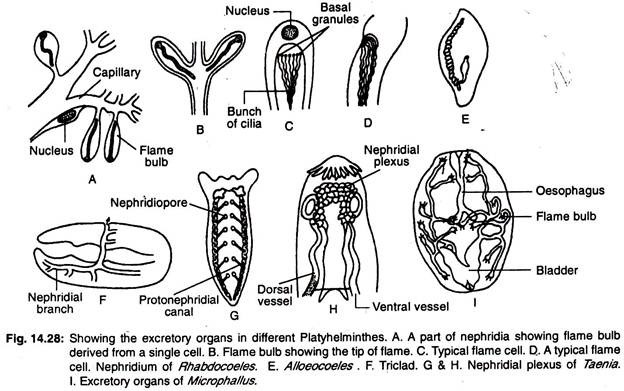
En los trematodos monogénicos hay dos conductos excretores longitudinales y los dos conductos se abren por separado en la superficie dorsal. Los conductos se dilatan para formar sacos excretores en sus extremos terminales.
En otros trematodos los conductos excretores están conectados posteriormente entre sí por un conducto transversal y se abren al exterior por una única abertura mediana. El conducto transversal dializa y en algunos casos forma una vesícula excretora. Los dos conductos longitudinales pueden unirse entre sí en la región posterior y pueden tener una sola abertura.
En los cestodos hay cuatro versículos excretores longitudinales. Los vasos excretores están conectados por un vaso transversal situado en el escólex para formar el plexo nefridinal (Fig. 14.28H) y se abren en una vesícula excretora contráctil situada en el último proglótido.
En los turbelarios de agua dulce hay numerosos protonefridios y un gran número de nefridioporos porque se impide la entrada de agua excesiva en el cuerpo.
Sistemas respiratorio y circulatorio de los platelmintos:
ADVERTENCIAS:
Estos dos sistemas están totalmente ausentes en el filo. En algunos trematodos se ha descrito la presencia de un sistema de tubos, denominado sistema linfático. Los tubos se consideran de funciones inciertas.
Sistema nervioso de los platelmintos:
El centro nervioso principal o cerebro está situado en la cabeza como un par de ganglios cerebrales. Del cerebro surgen varios cordones longitudinales ganglionados. De estos cordones longitudinales un par es el más conspicuo y el resto resulta insignificante. Entre los cordones longitudinales se producen numerosas conexiones transversales y todo el sistema nervioso adquiere un aspecto de escalera.
Los órganos sensoriales como los ocelos u ojos están presentes en los turbelarios y trematodos monogénicos. Son numerosos en los policlados y de dos a cuatro en los rabdocelos y los trematodos monogénicos. Los ojos están presentes como células pigmentarias o en forma de copas que contienen células pigmentarias y sensoriales. Los quimiorreceptores y tangorreceptores están ampliamente presentes y los estatocistos se dan en Acoela y en otras pocas formas.
Sistema reproductivo de Platyhelminthes:
A excepción de unos pocos Turbellaria y Trematoda, los platihelmintos son hermafroditas. Los órganos reproductores masculinos y femeninos de cada individuo están separados y se abren externamente por sus propios poros o por una apertura genital común. En algunos casos los gonoductos se abren en el tubo digestivo y las células sexuales se liberan por la boca.
ADVERTENCIAS:
En algunos platelmintos existe un poro femenino adicional o poro vaginal que sirve durante la cópula. Los gonoporos suelen estar en posición ventral, pero en ocasiones son dorsales. En las tenias el gonoporo es de posición lateral.
Los órganos reproductores masculinos consisten en testículos que en estado primitivo son numerosos y están dispersos. El número de testículos se reduce a uno o dos en muchos y en Acoela faltan gónadas y conductos definidos. Los vasa efferentia, cuando están presentes, corresponden al número de testículos presentes. En general, hay un par de vasa deferentia que se unen y se abren en el complicado aparato copulatorio.
El aparato copulatorio consiste en un cirro eversible o un pene protuberante armado con espinas o ganchos. Varias glándulas están asociadas a él. La vesícula, simple o emparejada, suele estar presente y sirve de depósito para almacenar los espermatozoides. Los órganos reproductores femeninos constan de uno o dos ovarios. Los oviductos cuando están emparejados se fusionan distalmente y forman un oviducto común que se abre en el aparato copulador.
El aparato copulatorio consta de un saco, la vesícula seminal o llamada bursa seminal o bursa copulatoria. Varias glándulas que contribuyen a la formación de la cáscara del huevo y a la producción de sustancias secretoras también están asociadas al aparato copulatorio. Un largo útero tubular o ramificado para acumular los huevos maduros forma una parte conspicua del aparato reproductor.
La gónada femenina de los platihelmintos es peculiar, ya que está claramente diferenciada en dos zonas, el ovario propiamente dicho y las glándulas vitelinas o vitelinas.
La yema nunca se incorpora a la estructura del huevo como en el caso de otros animales, sino que se produce como huevos abortivos que se incluyen dentro de la cápsula o cáscara del huevo para proporcionar alimento al embrión en desarrollo. En los cestodos los órganos reproductores se repiten en cada proglótido. En los proglótidos jóvenes los órganos son rudimentarios mientras que éstos están muy desarrollados en los grávidos.
Desarrollo de los platelmintos:
ADVERTENCIAS:
Aunque son hermafroditas, la práctica de la fecundación cruzada es la regla. La fecundación es interna y los huevos fecundados con cáscara que contienen embriones se desprenden al exterior. Los Acoela y los Policlados carecen de glándulas vitelinas y el desarrollo es directo. En las Policladas se produce una larva de natación libre que se supone que prefigura el Trocóforo.
La larva se llama larva de Muller o Protrochula (Fig. 14.29). Es de forma ovalada y lleva ocho brazos prominentes que están rodeados de largos cilios que forman una banda continua. La superficie general del cuerpo está cubierta de pequeños cilios.
La abertura bucal está situada en la línea media ventral y existen tres ojos en la parte anterior de la superficie dorsal. Durante el desarrollo los brazos ciliados son absorbidos. En otros grupos el desarrollo embrionario está muy modificado y complicado. 
Consideraciones filogenéticas de los platelmintos:
No existe una opinión unánime entre los zoólogos sobre el origen de los platelmintos. Hay varios puntos de vista para el origen de los platelmintos.
1. Lang (1881) propuso la teoría del ctenóforo-policládica. Esta teoría afirma que los turbelarios policládicos podrían haber evolucionado a partir de los ctenóforos a través de Platyctenea. La larva de Muller de los policlados tiene algunas semejanzas con los ctenóforos.
Los rasgos de semejanza son:
ADVERTENCIAS:
(i) Similitudes de las simetrías entre los ctenóforos y la larva.
(ii) Las 8 crestas cilíndricas ectodérmicas de la larva de Muller pueden compararse con las crestas meridionales de los ctenóforos.
(iii) El estatocisto de los turbelarios puede compararse con los ctenóforos.
Esta teoría no goza de mucho apoyo entre los zoólogos.
2. La teoría de la larva de la plánula afirma que los turbelarios tienen cierto parecido con la larva de la plánula de los cnidarios. Así, muchos zoólogos como Hyman (1951), Jagersten (1955) y Hadzi (1963), consideran que los platelmintos han evolucionado a partir de un tronco ancestral parecido a la plánula y que Acoela es el grupo más primitivo entre los platelmintos (Fig. 14.27A).
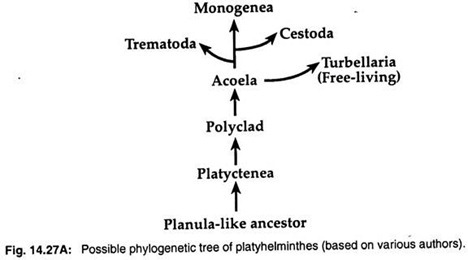
Los datos moleculares y los análisis cladísticos han sugerido un origen monofilético para los tres grupos parasitarios. Las tres clases parasitarias -Trematoda, Monogenea y Cestoda- han evolucionado a partir de turbelarios de vida libre.
3. De nuevo la teoría del enterocoel afirma que todos los animales bilaterales son básicamente coelomados, y los platelmintos acoelomados han evolucionado a partir de ancestros coelomados por pérdida de la cavidad de forma secundaria. Esta teoría no es aceptable para los zoólogos porque es difícil explicar algunos aspectos, como el cambio de la condición de coelomado a acoelomado y de la simetría bilateral a la radial.