ADVERTISEMENTS:
W tym artykule będziemy dyskutować o Platyhelminthes:- 1. Habitat i Habitat Platyhelminthes 2. Struktura Platyhelminthes 3. Narządy przylegania 4. Ściana ciała 5. Układ pokarmowy 6. Układ wydalniczy 7. Układ oddechowy 8. Układ nerwowy 9. Układ rozrodczy 10. Rozwój 11. Rozważania filogenetyczne.
- Habitat i środowisko życia platyhelminthes:
- Struktura Platyhelminthes:
- Organy przylegania lub ssawki u Platyhelminthes:
- Ściana ciała Platyhelminthes:
- Układ pokarmowy platyhelminthes:
- System wydalniczy platyhelminthes:
- Układy oddechowy i krążenia u Platyhelminthes:
- Układ nerwowy u Platyhelminthes:
- Układ rozrodczy Platyhelminthes:
- Rozwój Platyhelminthes:
- Phylogenetic Considerations of Platyhelminthes:
Habitat i środowisko życia platyhelminthes:
Platyhelminthes są w większości ekto lub endopasożytnicze, a nieliczne są wolno żyjące. Wolno żyjące należą do klasy Turbellaria i żyją w wodach słodkich, stawach, jeziorach, strumieniach i źródłach. Niektóre z nich występują na wybrzeżach w regionach tropikalnych i subtropikalnych. Trematoda i cestoda są pasożytami całkowitymi. W stadium dorosłym pasożytują na kręgowcach, a w stadium larwalnym występują jako pasożyty zwierząt bezkręgowych.
Struktura Platyhelminthes:
WSKAZANIA:
Członkowie tego azylu mają zwykle wydłużony wygląd. Polyclads są szerokie liści-jak, podczas gdy tasiemce są płaskie i wstążka-jak. Kontur ciała jest prosta w ogóle, ale niektóre trematodes oferują bizzare konturu. Ciało tasiemców składa się z wielu kwadratowych lub prostokątnych segmentów zwanych proglotydami.
Przedni proglotyd jest najmniejszy, a tylny największy. Oznacza to, że wielkość proglotydu wzrasta w kierunku przednio-tylnym. Obecność proglotydów nadaje tasiemcom cechę segmentacji. Pewien rodzaj pseudometameryzmu spotykany jest u Procerodes lobata, u którego niektóre narządy wewnętrzne powtarzają się.
Przednie i tylne końce, jak również powierzchnie grzbietowe i brzuszne są łatwo rozpoznawalne. Często przedni koniec jest oddzielony od reszty ciała przez obecność „głowy”, po której następuje zwężona „szyja”.
W niektórych formach brak jest wyraźnej głowy, ale przedni koniec może być wykryty przez narządy zmysłów lub przez jego ruch skierowany do przodu podczas lokomocji. Na powierzchni brzusznej znajdują się otwory gębowe i genitalne, jeśli są obecne.
POGLĄDY:
Rozmiary platyhelminthes wahają się od mikroskopijnych do ekstremalnie wydłużonych form o długości 10-15 metrów (tasiemce). Większość członków jest małych do umiarkowanych rozmiarów. Platyhelminthes są ogólnie bezbarwne lub białe, formy wolno żyjące są białe, brązowe, szare lub czarne. Niektóre poliklady i planarie lądowe mają jaskrawe kolory ułożone we wzory.
Organy przylegania lub ssawki u Platyhelminthes:
Platyhelminthes posiadają różnorodne narządy przylegania i przyczepiania. Acetabulum lub narząd ssący w postaci „przyssawki” jest bardzo rozpowszechniony u dorosłych robaków płaskich. U motyli występują dwa przyssawki na brzusznej stronie ciała. Jeden z nich znajduje się w przedniej części ciała, a jego położenie jest mniej więcej stałe.
Natomiast druga, zwana tylną przyssawką, nie ma stałego położenia. U Paramphistomum przyssawka ta jest położona najbardziej z tyłu, u Echinostoma i Fasciola jest przesunięta bardziej do przodu.
W tasiemcach narządy przylepne występują w postaci rowków lub miseczek i znajdują się na końcu głowowym. Często w pobliżu tych rowków lub w odwracalnej komorze policzkowej znajdują się haczyki ułatwiające zakotwiczenie. Przyssawki występują również u niektórych wolno żyjących planarii.
Ściana ciała Platyhelminthes:
Platyhelminthes brak egzo- lub endoszkieletu i jako takie ciało jest miękkie. Naskórek jest jednowarstwowy, a w niektórych przypadkach jest syncytialny. Naskórek może być w całości lub w części pokryty rzęskami. Warstwa podnaskórkowa składa się z mięśniówki o włóknach okrężnych, podłużnych i skośnych. Włókna mięśniowe są gładkie. Wszystkie przestrzenie między organami wypełnione są komórkami wypełniającymi, zwanymi parenchymą.
Części twarde u Platyhelminthes są haki i kolce. Wielu wcześniejszych pracowników uważało, że ciało monogeneans, digeneans i cestodes jest pokryte „kutykulą”.
Non-ciliation w tych formach daje fałszywe wrażenie kutykuli w mikroskopii świetlnej. Ale badania mikroskopem elektronowym ujawniły, że zewnętrzną warstwą ciała jest epiderma. Komórki w tej warstwie zawierają mikrochondria i pozostają w ciągłości z komórkami podstawowymi.
Układ pokarmowy platyhelminthes:
Układ trawienny nie występuje u turbelarii z rzędu Acoela i u cestodów. U Turbellaria i Trematoda układ pokarmowy jest reprezentowany przez jamę gębową, gardło i jelito, które kończy się ślepo, i jako takie całe rozmieszczenie narządów trawiennych przypomina ściśle układ pokarmowy antozoanów i ctenoforów.
Usta w stanie prymitywnym znajdują się mniej więcej w połowie linii brzusznej, ale u wielu płazińców pozycja ta jest przesunięta do przodu wzdłuż linii brzusznej.
WSKAZANIA:
Gardło jest nieobecne u endopasożytniczych rabdoków. Gardło ma charakter stomodalny i jest silnie umięśnioną rurką. To pokazuje zróżnicowanie w azylu. Jelito wykazuje duże zróżnicowanie pod względem kształtu. Może być prostym workiem lub mieć skomplikowane rozgałęzienia i podgałęzienia. Otwory odbytowe są rzadko obecne.
System wydalniczy platyhelminthes:
Układ wydalniczy platyhelminthes składa się głównie z podłużnych naczyń wodnych lub podłużnych kanałów wydalniczych posiadających szereg wyspecjalizowanych komórek wydalniczych, zwanych komórkami płomieniowymi lub cebulkami płomieniowymi, zwanych zbiorczo protonephridia. Rozmieszczenie naczyń wodnych wewnątrz ciała daje szeroki zakres zmienności.
W triclads znajdują się dwa podłużne naczynia (kanały), które otwierają się na zewnątrz licznymi porami (ryc. 14.28G). W rhabdocoelach występują dwa naczynia boczne lub jedno przyśrodkowe (ryc. 14.28E). Naczynie przyśrodkowe otwiera się na zewnątrz przez pojedynczy por znajdujący się na tylnym końcu ciała.
W formach, u których występują dwa naczynia, otwory znajdują się albo po stronie brzusznej, albo w gardzieli. W niektórych formach oba naczynia łączą się w tylnej części i otwierają się na zewnątrz jednym środkowym otworem. Główny kanał nefrydialny jest szeroko rozgałęziony i oprócz nefrydioforu występują w nim dodatkowe pory wydalnicze (ryc. 14.28F).
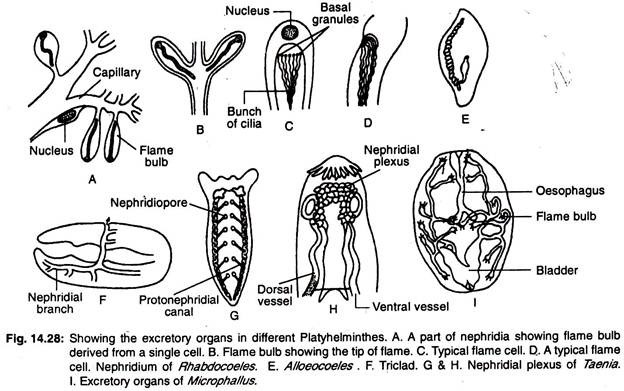
W monogenicznych trematodach występują dwa podłużne przewody wydalnicze i oba przewody otwierają się oddzielnie na powierzchni grzbietowej. Przewody rozszerzają się tworząc worki wydalnicze na swoich końcowych końcach.
W innych trematodach przewody wydalnicze są połączone z tyłu ze sobą przewodem poprzecznym i otwierają się na zewnątrz pojedynczym otworem przyśrodkowym. Przewód poprzeczny dializuje i w niektórych przypadkach tworzy pęcherzyk wydalniczy. Dwa podłużne kanały mogą łączyć się ze sobą w tylnej części i mogą mieć pojedynczy otwór.
W cestodach występują cztery podłużne wersaliki wydalnicze. Naczynia wydalnicze połączone są poprzecznym naczyniem znajdującym się w scolexie, tworząc splot nefrydynalny (ryc. 14.28H) i otwierają się w kurczliwym pęcherzyku wydalniczym znajdującym się w ostatnim proglotydzie.
Nieliczne protonefridia i duża liczba nefridioporów występują u turbulistów słodkowodnych, ponieważ zapobiega się napływowi nadmiaru wody do organizmu.
Układy oddechowy i krążenia u Platyhelminthes:
UWAGI:
Te dwa układy są całkowicie nieobecne w tym azylu. W niektórych trematodach opisano obecność systemu rurek, zwanego układem limfatycznym. Rurki te uważane są za pełniące niepewne funkcje.
Układ nerwowy u Platyhelminthes:
Główny ośrodek nerwowy lub mózg znajduje się w głowie w postaci pary zwojów mózgowych. Kilka ganglionated podłużne sznury powstają z mózgu. Z tych podłużnych sznurów jedna para staje się najbardziej widoczna, a reszta staje się nieistotna. Pomiędzy sznurkami podłużnymi powstają liczne połączenia poprzeczne, a cały układ nerwowy nabiera wyglądu drabiniastego.
Organy zmysłów w postaci oczodołów lub oczu występują u turkuciów i monogenicznych trematodów. Są one liczne w polyclads i dwa do czterech w Rhabdocoels i monogeniczne trematodes. Oczy występują jako komórki pigmentowe lub w postaci miseczek zawierających komórki pigmentowe i czuciowe. Chemo i tangoreceptory są obecne powszechnie, a statocysty występują u Acoela i kilku innych form.
Układ rozrodczy Platyhelminthes:
Z wyjątkiem nielicznych Turbellaria i Trematoda, platyhelminthes są hermafrodytami. Męskie i żeńskie narządy rozrodcze u każdego osobnika są oddzielne i otwierają się zewnętrznie przez ich własne pory lub przez wspólny otwór płciowy. W niektórych przypadkach gonodukty otwierają się do przewodu pokarmowego i komórki płciowe są uwalniane przez jamę ustną.
WSKAZANIA:
W niektórych płazińcach występuje dodatkowy por samicy lub por pochwowy, który służy podczas kopulacji. Gonopory są zwykle ułożone brzusznie, ale czasami bywają grzbietowe. U tasiemców gonopory znajdują się w pozycji bocznej.
Męskie narządy rozrodcze składają się z jąder, które w stanie pierwotnym są liczne i rozproszone. W wielu przypadkach liczba jąder jest zredukowana do jednego lub dwóch, a u Acoela brak jest wyraźnych gonad i przewodów. Vasa efferentia, jeśli są obecne, odpowiadają liczbie obecnych jąder. Ogólnie rzecz biorąc, istnieje para vasa deferentia, które łączą się i otwierają w skomplikowany aparat kopulacyjny.
Przyrząd kopulacyjny składa się z odwracalnego prącia lub wystającego prącia uzbrojonego w kolce lub haki. Związane są z nim różne gruczoły. Pęcherzyk nasienny w stanie pojedynczym lub sparowanym jest często obecny i działa jako zbiornik do przechowywania plemników. Narządy rozrodcze samicy składają się z jednego lub dwóch jajników. Jajowody, gdy są sparowane, łączą się dystalnie i tworzą wspólny jajowód, który otwiera się do aparatu kopulacyjnego.
Aparat kopulacyjny składa się z woreczka, pęcherzyka nasiennego lub zwanego bursą nasienną lub bursą kopulacyjną. Z aparatem kopulacyjnym związane są również różne gruczoły, które pomagają w tworzeniu skorupek jajowych i produkcji substancji wydzielniczych. Długa rurkowata lub rozgałęziona macica, służąca do gromadzenia dojrzałych jaj, stanowi widoczną część układu rozrodczego.
Gonady żeńskie u platyhelminthes są osobliwe, ponieważ są wyraźnie zróżnicowane na dwie strefy, jajnik właściwy i gruczoły żółtkowe lub szklistkowe.
Żółtko nigdy nie jest włączone do struktury jaja, jak w przypadku innych zwierząt, lecz jest produkowane jako jajo poronne, które ma być włączone do wnętrza kapsuły jajowej lub skorupy w celu zapewnienia pożywienia dla rozwijającego się zarodka. U cestodów narządy rozrodcze powtarzają się w każdym proglotydku. U młodych proglotydów narządy te są rudymentarne, podczas gdy u jajorodnych są one silnie rozwinięte.
Rozwój Platyhelminthes:
ZACHOWANIA:
Chociaż hermafrodytyczne, praktyka zapłodnienia krzyżowego jest regułą. Zapłodnienie jest wewnętrzne, a zapłodnione, pokryte skorupkami jaja zawierające embriony są wydalane na zewnątrz. U Acoeli i Polycladów brak gruczołów żółtkowych, a rozwój jest bezpośredni. U Polycladów powstaje swobodnie pływająca larwa, która ma być zapowiedzią trochofora.
Larwa ta nazywana jest larwą Mullera lub Protrochula (ryc. 14.29). Ma ona kształt owalny i posiada osiem wydatnych ramion, które są obrzeżone długimi rzęskami tworzącymi jedno ciągłe pasmo. Ogólna powierzchnia ciała pokryta jest drobnymi rzęskami.
Phylogenetic Considerations of Platyhelminthes:
Wśród zoologów nie ma jednomyślnego poglądu co do pochodzenia płazińców. Istnieje kilka poglądów na pochodzenie płazińców.
1. Lang (1881) zaproponował teorię ctenophore- polyclad. Teoria ta głosi, że wielopłaskonose turbulary mogły powstać z ctenoforów poprzez Platyctenea. Larwa Mullera z polyclad wykazuje pewne podobieństwa z ctenoforami.
Cechy podobieństwa to:
PODOBIEŃSTWA:
(i) Podobieństwa symetrii między ctenoforami i larwą.
(ii) 8 ektodermalnych grzbietów cilialnych larwy Mullera można porównać z grzebieniami meridialnymi kenoforów.
(iii) Statocysta turbulistów może być porównana z ctenoforami.
Ta teoria nie cieszy się dużym poparciem wśród zoologów.
2. teoria larwy planulacyjnej stwierdza, że turbularze wykazują pewne podobieństwa do larw planulacyjnych knidarii. Tak więc wielu zoologów, jak Hyman (1951), Jagersten (1955) i Hadzi (1963), uważa, że płazińce wyewoluowały z planulopodobnego przodka, a Acoela jest najbardziej prymitywną grupą wśród platyhelminthów (ryc. 14.27A).
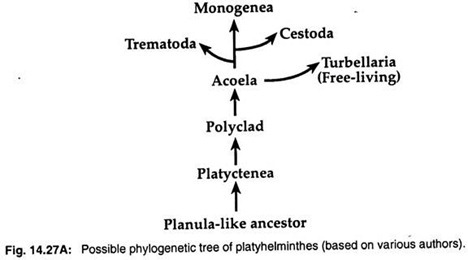
Dane molekularne i analizy kladystyczne sugerują monofiletyczne pochodzenie trzech grup pasożytniczych. Trzy pasożytnicze klasy – Trematoda, Monogenea i Cestoda – wyewoluowały z wolno żyjących turbanów.
3. Ponownie teoria enterocoel stwierdza, że wszystkie zwierzęta obojnacze są w zasadzie koelomate, a płazińce akoelomate wyewoluowały z przodków koelomate przez wtórną utratę jamy. Teoria ta jest nie do przyjęcia przez zoologów, ponieważ trudno jest wyjaśnić niektóre aspekty, takie jak przejście od stanu koelomatu do akoelomatu i od symetrii dwustronnej do promienistej.