Learning Outcomes
- Identify historical and potential causes of high extinction rates
地球上、あるいは任意の地理領域の種の数は、継続的に進行中の二つの進化プロセスの平衡の結果です:種分化と絶滅です。 どちらも大進化の自然な「誕生」と「死」のプロセスである。 種分化の速度が絶滅の速度を上回り始めると、種の数は増加し、同様に、絶滅の速度が種分化の速度を上回り始めると、種の数は減少する。 地球の歴史を通じて、この2つのプロセスは変動しており、時には化石記録に反映されているように、地球上の種の数が劇的に変化することもあった(図1)
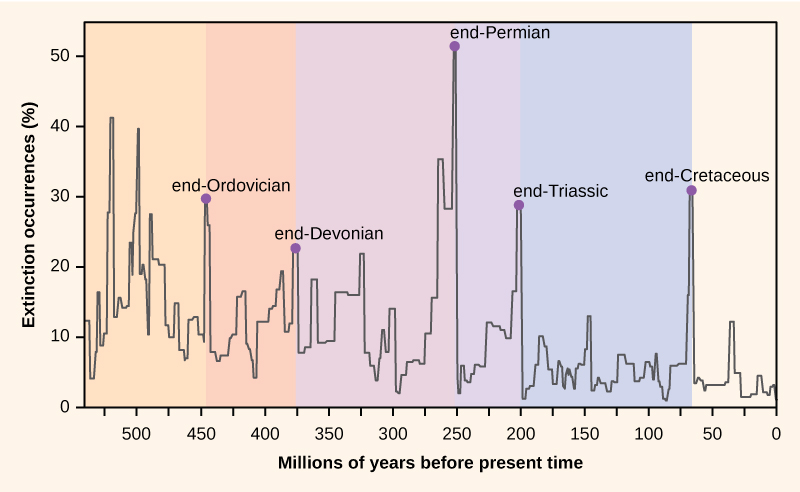
図1. 図1 化石記録に見られる絶滅の割合は、地球の歴史を通じて変化してきた。 古生物学者は、化石記録の中で、生物多様性の突然の劇的な損失(化石記録から現存する種の半分以上が消えている)を示すと思われる5つの地層を特定している。 これらは大量絶滅と呼ばれています。 しかし、この5つの大量絶滅は、最も多くの研究を集めている。 この5つの大量絶滅は、新生代(5億4200万年前以降)の連続した大規模な絶滅現象の中で、最も極端な5つの現象に過ぎないという主張ができる。 ほとんどの場合、仮説とされる原因はまだ論争の的となっているが、しかし、最も最近の出来事は明らかであると思われる。
Recorded Mass Extinctions
大量絶滅の化石記録は地史の期間を定める基礎となったので、一般に地史の間の移行点で発生することがある。 ある時代から別の時代への化石の推移は、種の劇的な喪失と新しい種の漸進的な起源を反映している。 このような変遷は、岩石の地層に見ることができる。 表1は、5つの大量絶滅に関するデータである。 大量絶滅
オルドビス紀-シルル紀の絶滅現象は、記録上最初の大量絶滅であり、2番目に大きい絶滅である。 この時期、海洋生物種(海洋以外に生息する種はほとんどない)の約85%が絶滅した。 その原因については、氷河期とその後の温暖化が主な仮説とされている。 しかし、この絶滅現象は、実際には100万年程度の時間を隔てた2つの絶滅現象から構成されている。 1回目は冷却によるもので、2回目はその後の温暖化によるものである。 気候の変化は気温と海水準に影響を与えた。 オルドビス紀-シルル紀の絶滅は、近くの超新星によるガンマ線バーストが原因である可能性を指摘する研究者もいます。 ガンマ線バーストは、地球のオゾン層を破壊し、太陽からの強い紫外線を地表に到達させた可能性があり、当時の気候変動を説明するものと考えられています。 ただし、この仮説はあくまで推測であり、地球の歴史に地球外生命体がどのような影響を及ぼしたかは、現在も研究が続けられている。 大量絶滅後の生物多様性の回復には、場所によって500万年から2,000万年かかった。 そのほとんどは海洋生物に影響を与え、地上に生息する植物や動物にはあまり影響を与えなかったようである。 この絶滅の原因はよく分かっていない。
ペルム紀末の絶滅は、生命の歴史の中で最大のものであった。 実際、この絶滅イベントで地球はほとんど生命がいなくなったという議論がなされるかもしれない。 推定では、全海洋種の96パーセント、全陸上種の70パーセントが失われたとされている。 例えば、オルドビス紀・シルル紀の絶滅を生き延びた三葉虫が絶滅したのもこの時である。 この大量絶滅の原因は明らかではないが、火山活動が広範囲に拡大し、地球温暖化が暴走したことが有力視されている。 海はほとんど無酸素状態になり、海洋生物は窒息死してしまった。 地球上の四肢動物の多様性は、ペルム紀末の絶滅の後、3000万年かけて回復した。 ペルム紀の絶滅は、地球の生物多様性の構成と進化の過程を劇的に変えた。
三畳紀-ジュラ紀絶滅イベントの原因は明確ではなく、研究者は気候変動、小惑星の衝突、火山噴火などの仮説を唱えている。 この絶滅イベントは超大陸パンゲアが分裂する直前に発生したが、最近の研究では、絶滅は三畳紀を通じてもっと徐々に発生したのではないかと考えられている。
白亜紀末の絶滅イベントの原因は、最もよく理解されているものである。 この絶滅の原因は、現在のユカタン半島の沖合に大きな隕石(小惑星)が激突した結果であると、現在では理解されている。 この仮説は1980年に初めて提唱され、白亜紀と古第三紀の境界を示す岩石層中のイリジウム(隕石から一定の割合で大気中に侵入するが、地表には存在しない)の濃度が急激に上昇したことに基づく過激な説明であった(図2)。 この境界は、化石中の恐竜だけでなく、他の多くの分類群も消滅したことを意味する。 イリジウムスパイクを発見した研究者は、この時期に堆積物の堆積が遅くなったのではなく、宇宙から(大きな小惑星の形で)イリジウムが大気中に急激に流入したと解釈した。 これは過激な説明だったが、1991年に適切な年齢と大きさの衝突クレーターが報告されたことで、この仮説の信憑性が高まった。 現在では、この説を裏付ける地質学的な証拠が数多く見つかっている。 白亜紀末の絶滅後の生物多様性の回復時間は、地質学的時間ではペルム紀末の絶滅よりも短く、1000万年のオーダーである。
もう一つの可能性は、おそらくユカタン小惑星の衝突と同時に、ユカタン小惑星の衝突とほぼ同じ時期、約6600万年前の白亜紀の終わりに形成が始まった大規模な火山活動であった。 溶岩流は現在のインドの50%以上を覆っていた。 トラップ形成時の火山ガス、特に二酸化硫黄の放出は気候変動に寄与し、大量絶滅を誘発したと考えられる。
練習問題
1980年、ルイスとウォルター・アルバレス、フランク・アサロ、ヘレン・ミケルスは、世界中で、K-Pg境界の堆積層内のイリジウム濃度にスパイクがあることを発見しました。 このイリジウムスパイクは、K-Pg大量絶滅をもたらした小惑星の衝突によって引き起こされたというのが、研究者たちの仮説であった。 図2において、イリジウム層は光の帯です。
研究者はこの岩石サンプルにおいて、K-Pg境界の上と下にあるシダの胞子の相対的な存在量を測定しました。
- K-Pg境界より下では、いくつかの種からなるシダの胞子が豊富に見つかったが、上では見つからなかった。
- K-Pg境界より上で、いくつかの種からなるシダの胞子が豊富に見つかったが、下では見つからなかった。
- K-Pg境界の上でも下でもシダの胞子が豊富に見つかったが、境界の下では1種しか見つからず、境界の上では多くの種が見つかった。
- 境界の上でも下でも多くの種のシダの胞子が見つかったが、その総数は境界の下でより多かった。
更新世の絶滅
更新世の絶滅は、より少ない絶滅の一つで、最近のものである。 最終氷期の終わりに北米やユーラシアの大型の脊椎動物が絶滅したことはよく知られている。 この絶滅は、1万年から1万2千年前という比較的限られた期間に起こったようである。 北米では、マンモス(約4,000年前までカナダのウランゲル島に現存)、マストドン、ビーバー、ナマケモノ、ネコ科の動物、ラクダなど、非常に多くの動物が失われた。 1900年代初頭、科学者たちは、これらの大型動物が急速に絶滅したのは、乱獲が原因である可能性を初めて指摘した。 この仮説に関する研究は現在も続けられている。
一般に、更新世の絶滅の時期は、おそらく 4 万年前の古人類の到来と相関しており、これらの絶滅の主な競合仮説である気候変動現象とは無関係であった。 オーストラリアでは、人類が到来した直後の約4万〜5万年前から、有袋類のライオン、1トンの巨大ウォンバット、巨大カンガルーの数種が姿を消している。 北米では、1万〜1万2千年前にほぼすべての大型哺乳類が絶滅している。 現在残っているのは、クマ、ヘラジカ、ヘラジカ、クーガーなどの小型哺乳類だけです。 最後に、海洋上の多くの離島では、多くの種の絶滅が、人類の到来と同時に起こった。 すべての島に大型動物がいたわけではないが、大型動物がいた場合、絶滅に追い込まれることが多い。 マダガスカルは約2000年前に植民地化され、そこに生息していた大型哺乳類が絶滅してしまったのです。 ユーラシア大陸やアフリカ大陸ではこのようなパターンは見られませんが、狩猟採集民であった人類が最近になってやってきたわけでもありません。 ユーラシア大陸に人類がやってきたのは、数十万年前である。 このテーマは、現在も活発に研究され、仮説が立てられている分野である。 2397>
現代の絶滅
6 番目の、つまり完新世の大量絶滅は、これまで考えられていたよりも早く始まったようで、現代のホモサピエンスの破壊的な活動が大きな原因であることが明らかになりました。 完新世が始まって以来、人類の著作に記録されている個々の種の最近の絶滅が多数ある。 そのほとんどは、1500年代以降のヨーロッパ植民地の拡大と一致している。
初期の例としてよく知られているのは、ドードー鳥である。 モーリシャス(インド洋の島)の森に住んでいた鳩のような奇妙な鳥で、1662年ごろに絶滅した。 ドードーは船乗りに食肉として狩られ、人間を恐れずに近づいてくるので格好の獲物だった(ドードーは人間とは進化していない)。 また、ヨーロッパの船が島に持ち込んだ豚、ネズミ、犬もドードーの子供や卵を殺した。
1768年に絶滅したステラー海牛は、マナティーに近縁で、おそらくかつて北米の北西海岸に生息していた。 1741年にヨーロッパ人によって初めて発見されたステラー海牛は、肉や油のために乱獲された。 最後のウミウは1768年に殺された。 これは、海牛がヨーロッパ人と最初に接触してから種が絶滅するまでのわずか27年間にあたる!
1900 年以降、さまざまな種が絶滅した。 この種はかつて渡り鳥として北米の空を暗くしていたが、乱獲され、農地のための森林伐採による生息地の喪失に苦しんだ。
これらは、過去500年間に記録された絶滅のほんの一部にすぎません。 国際自然保護連合(IUCN)は、レッドリストと呼ばれる絶滅種や絶滅のおそれのある種のリストを作成しています。 このリストは完全ではありませんが、西暦1500年以降の脊椎動物の絶滅種380種が記載されており、そのうち86種は乱獲や乱獲によって絶滅しました。
現在の絶滅率の推定
絶滅率の推定は、ほとんどの絶滅がおそらく観察なしで起こっているという事実が妨げとなっています。 鳥類や哺乳類の絶滅は、特に狩猟やその他の方法で利用されていた場合、人間の目に留まる可能性が高い。 しかし、人間にとってあまり興味のない(必ずしも価値が低いわけではない)生物や、記述されていない生物もたくさんある。
バックグラウンドの絶滅率は、年間100万種につき1種程度(E/MSY)と推定されている。 たとえば、約1000万種が存在すると仮定すると、毎年10種が絶滅すると予想される(各年は年間1000万種に相当する)。
現代のある絶滅率の推定では、1500年以降の文書記録における絶滅を使用している。 鳥類だけで、この方法は26E/MSYという推定値を得た。 しかし、この値は3つの理由から過小評価である可能性がある。 第一に、多くの種はその時代のかなり後期まで記述されなかったであろうから、その損失は気づかれなかったであろう。 第二に、最近絶滅した脊椎動物の種は、現在、骨格から絶滅種が記載されるようになっているため、その数は増加している。 第三に、自然保護論者が名前をつけたがらないが、おそらくすでに絶滅している種もある。 これらの要因を考慮すると、推定絶滅率は100E/MSYに近くなる。 今世紀末までに予測される絶滅率は1500E/MSYである。

図3。 生息地の広さに応じて存在する種の数が増加することが研究で示されている。 (credit: modification of work by Adam B. Smith)
現在における絶滅率を推定するための第2のアプローチは、森林面積の損失を測定し、種と面積の関係を把握することによって種の損失と生息地の損失を相関させることである。 種-面積関係とは、調査した面積を増やしたときに新しい種が見られる割合のことである。 島の大きさが大きくなるにつれて、存在する種の数が増えるという研究結果が出ています。 この現象は、熱帯雨林に囲まれたベネズエラの山頂テプイのような、他の島のような生息環境でも同様に起こることが示されている。 この関係を逆転させると、生息地の面積が減少すれば、そこに生息する種の数も減少することになる。 生息地の消失と種面積の関係から絶滅率を推定すると、約90%の生息地が消失した場合、50%の種が絶滅すると予測されている。 種-面積の推定により、種の絶滅率は約1000E/MSY以上と計算されている。 一般に、実際の観測ではこれほどの損失は見られず、絶滅が遅れているとの指摘がなされている。 また、最近の研究では、種の喪失を推定する際の種-面積の関係の適用性に疑問が投げかけられている。 この研究では、種と面積の関係は絶滅率の過大評価につながると論じている。 より適切な関係は、固有種-面積の関係であろう。 この方法を用いれば、今後100年間の推定値は500E/MSY程度まで下がるだろう。
Try It
Contribute!
このページを改善するもっと知る
。