ADVERTISEMENTS:
この記事では扁形動物について説明します: – 1.扁形動物の生息地と生息地 2.扁形動物が生息している場所 3.扁形動物が生息している場所 4.扁形動物が生息している場所 5.扁形動物の生息地と生息地 6.扁形動物の生息地 7.扁形動物の生息地 9. 扁形動物門の構造 3. 付着器官 4. 体壁 5. 消化器系 6. 排泄器官 7. 呼吸器系 8. 神経系 9. 生殖器系 10. 発生 11. 系統学的考察
扁形動物類の生息・生育環境:
扁形動物類は、ほとんどが外生または内生寄生で、自由生は少ない。 自由生活型のものはTurbellariaに属し、淡水、池、湖、小川、泉などに生息しています。 また、熱帯・亜熱帯の海岸に生息するものもある。 トレマトーダとセストーダは完全な寄生虫である。 成虫期には脊椎動物に寄生し、幼虫期には無脊椎動物に寄生する。
扁形動物門の構造。
扁形動物門の構造:
この門の仲間は通常、細長い外見をしています。 ポリクラッドは広い葉のようで、サナダムシは扁平でリボンのようです。 体の輪郭は一般に単純であるが、トレマトードの中には奇妙な輪郭を持つものもある。 サナダムシの体は、プログロティッドと呼ばれる四角形または長方形のセグメントが多数集まってできている。
前方の突起が最も小さく、後方の突起が最も大きい。 つまり、前後方向に大きくなっています。 吻節があることで、サナダムシは分節化した状態になる。 Procerodes lobataでは、内臓の一部が繰り返される、ある種の擬似同型体である。
前端、後端、背面、腹面は容易に識別できる。 前端は「頭」と「頸」があり、他の部分と区別できることが多い。
明確な頭部がないものもあるが、前端は感覚器官によって、あるいは運動時に前方に向けられる動きによって感知できる。 腹面には口と生殖器がある場合は、それがある。
扁形動物には、微小なものから、体長10~15mの極端に細長いもの(サナダムシ)までいる。 ほとんどの仲間は小型から中型の大きさである。 一般に扁形動物類は無色か白色で、自由行動するものは白色、茶色、灰色、黒色をしている。 多足類や陸生プラナリアには、鮮やかな色彩で模様を描くものもある。
扁形動物における付着器官と吸盤。
扁形動物にはさまざまな付着器官や吸盤がある。 扁形動物成虫では’吸盤’の形の付着器官や吸盤が非常に多く見られます。 フクラギ類では、腹側に2つの吸盤がある。 吸盤の1つは体の前側にあり、その位置は多かれ少なかれ固定されている。
一方、後吸盤と呼ばれるもう1つの吸盤は位置が一定していない。 Paramphistomumでは吸盤の位置が最も後方にあり、EchinostomaやFasciolaではより前方にずれている。
サナダムシでは、粘着器官は溝やカップの形で存在し、頭端に位置している。 この溝付近や頬部には固定を補助するための鉤が存在することが多い。 吸盤は自由生活するプラナリアの一部にも存在する。
扁形動物門の体壁。
扁形動物には外骨格や内骨格がなく、体は軟らかい。 表皮は単層で、場合によっては合着していることもあります。 表皮の一部または全部が繊毛化することもある。 表皮下層は、円形、縦長、斜めの繊維からなる筋層で構成されている。 筋繊維は平滑である。 臓器間の空間はすべて実質細胞と呼ばれるパッキング細胞で満たされている。
扁形動物門の硬い部分は鉤と棘である。 単弓類,掘脚類,条虫類は体が「クチクラ」に覆われていると考える先達が多い。
これらの形態では繊毛がないため、光学顕微鏡で見るとクチクラのような印象を受ける。 しかし、電子顕微鏡の研究によって、体の外側の層は表皮であることが明らかになっている。 この層の細胞はマイクロコンドリアを含み、下層の細胞と連続したままである。
扁形動物類の消化器官。
ターボラ目のアコエラや条虫類には消化器官はない。 ターベラリアとトレマトーダでは消化器系は口、咽頭、盲腸で終わり、そのため消化器官の全体の配置はアントゾアンや有櫛動物に酷似している。
口は原始的な状態では腹線のほぼ中央に位置しているが、多くの扁形動物では腹線に沿って前方にずれている。
補足:
内寄生性のラブドコアでは口は存在しない。 咽頭は気孔を持ち,強い筋肉質の管である。 門によって変異が見られる。 腸の形態は様々である。 単純な袋状のものから、複雑な枝分かれをするものまである。 肛門開口部はほとんど存在しない。
扁形動物門の排泄器官。
扁形動物の排泄系は、主に縦長の水管または縦長の排泄管からなり、火炎細胞または火炎球と呼ばれる多数の特殊な排泄細胞を持ち、これらをまとめて原口と呼びます。 体内の水管の配置はバリエーションに富んでいる。
三葉虫では、2本の縦管(運河)があり、多数の孔で外部に開かれている(図14.28G)。 ラブドクエルには2本の横管または1本の中央管がある(図14.28E)。 中央の血管は体の後端にある1つの孔で外部に開口する。
2本の血管があるものは、開口部が腹側か咽頭部にある。 2本の管が後方で合流し、中央の1つの開口部で外部に開口するものもある。 主なネフリ管は大きく分岐し、ネフリジオポアのほかに付属の排泄孔がある(図14.28F)。
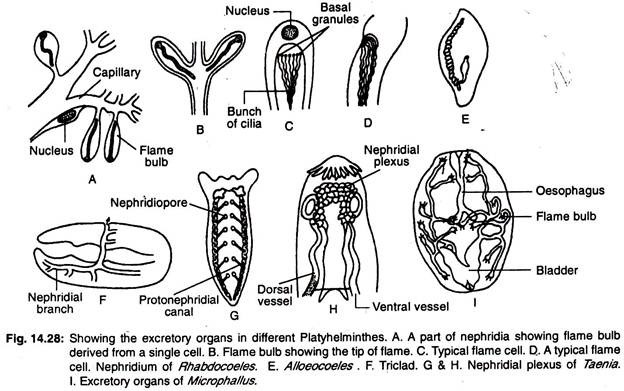
単弓類では縦長の排泄管が2本あり、2本の管は背面で別々に開口している。 管は拡張し、末端には排泄嚢が形成される。
他の振戦動物では、排泄管は横管で互いに後方でつながり、中央の1つの開口部で外部に開口している。 横管は透析し、場合によっては排泄胞を形成する。 2本の縦管は後方領域で互いに合流し、1つの開口部を持つこともある。
条虫では4本の縦走排泄胞がある。 これらの排泄管は鞘腔内にある横管で連結されて腎叢を形成し(図14.28H)、最後の前鰓孔にある収縮性の排泄小胞に開口している。
淡水産のターベラスでは、体内への過剰な水の流入を防ぐため、多数の原虫鞘と多数のネフリジオポアが存在します。
扁形動物の呼吸・循環器系:
注意:
この2つのシステムは、この門では全く存在しません。 いくつかの線虫では、リンパ系と呼ばれる管のシステムの存在が報告されています。 この管は機能不明とされている。
扁形動物門の神経系。
主な神経中枢(脳)は、一対の大脳神経節として頭部に位置する。 脳からはいくつかの神経節をもつ縦索が出ています。 これらの縦索のうち、1対が最も顕著になり、残りは重要でなくなる。 縦索の間には多数の横索が発生し、神経系全体が梯子状になる。
眼球などの感覚器はターベリアンや単発性振戦動物に存在する。 多足類では多数,ラブドコエルや単発性振戦動物では2〜4個である。 眼は色素細胞として、あるいは色素細胞や感覚細胞を含むカップの形で存在する。 ケモやタンゴレセプターは広く存在し、スタトシストはアコエラや他の形態ではほとんど見られない。
扁形動物類の生殖器官:
一部のターベラリアとトレマトーダを除いて、扁形動物類は雌雄同体である。 各個体の雄生殖器と雌生殖器は別々で,それぞれの孔で外部に開口するか,共通の生殖器開口部で開口する。 生殖管が消化管に開口し、口から性細胞が遊離する場合もある。
注意:
いくつかの扁形動物では、交尾の際に機能する余分な雌孔または膣孔が存在します。 生殖孔は通常腹側にあるが、背側にある場合もある。 サナダムシでは、生殖孔は側方にある。
雄の生殖器官は精巣からなり、原始的な状態では多数、散在している。 精巣の数は1〜2個に減少するものが多く、アコエラでは明確な生殖腺や管はない。 精巣がある場合、その数は精巣の数に対応する。 一般に精管は1対で結合し、複雑な交尾器官へと開く。
交接器は、棘や鉤状の突起で武装した、逆さになった円柱または突出した陰茎で構成されています。 また、様々な分泌腺が付随しています。 精嚢は単一または対の状態で存在することが多く,精子を貯蔵する貯蔵庫として機能する。 雌の生殖器官は1個または2個の卵巣からなる。 卵管は対になっている場合、遠位で融合し、共通の卵管を形成し、交尾器に開口する。
交接器は、精嚢または精嚢あるいは交接包と呼ばれる袋で構成されている。 卵殻の形成や分泌物の生成に役立つ様々な腺も、交接器に付随しています。 熟した卵を蓄積するための長い管状または分岐した子宮は、生殖器系の顕著な部分を形成している。
扁形動物の雌の生殖腺は、卵巣と卵黄腺の2つにはっきりと分かれているのが特徴です。
卵黄は他の動物のように卵の構造に組み込まれることはなく、卵嚢や殻の中に含まれ、発達中の胚に食物を提供するための休止卵として作られる。 絛虫類では、生殖器官はそれぞれの胞子嚢の中で繰り返される。 若い胞子虫では器官は初歩的であるが、妊娠中の胞子虫では器官は高度に発達している。
扁形動物類の発生。
注意事項:
両性具有であるが、交配を行うのが原則である。 受精は内部で行われ、受精した胚を含む殻付き卵は外部に排出される。 アコエラとポリクラッドには卵黄腺がなく、発生は直接的である。 ポリクラッドでは自由遊泳する幼生が作られるが、これはトロコフォアの前兆とされる。
この幼虫はミュラー幼虫またはプロトロクラと呼ばれる(図14.29)。 形は楕円形で、8本の腕があり、その腕には長い繊毛が一続きの帯状に付いています。 体表は小さな繊毛で覆われている。
口は腹部中央にあり、背面前方には3個の眼がある。 発生過程で繊毛腕は吸収される。 他の群では胚発生が大きく変化し、複雑になっています。 
扁形動物門の系統的考察。
扁形虫の起源については、動物学者の間で統一した見解はない。 扁形動物の起源についてはいくつかの見解がある。
1. Lang(1881)はctenophore-polyclad説を提唱した。 この説は、多足類タービュラリアンがctenophoreからPlatycteneaを経て進化した可能性があるとするものである。 この説では、Muller’s larvaがctenophoresと似ていることから、polycladの幼生がctenophoresの幼生に似ているとしている。
似ている点は以下の通り。
注意点:
(i)クテノフォアと幼生の対称性が似ていること。
(ii)ミュラーズ幼生の8つの外皮繊毛隆起は、有櫛動物の子午面櫛形板と比較できる。
(iii)タービリアンの静止胞子は有櫛動物に例えることができます。
この説は動物学者の間ではあまり支持されていない。
2.扁平幼生説は、刺胞動物の扁平幼生に類似しているとするもので、扁平幼生が刺胞動物の扁平幼生に似ているとするものである。 そのため,Hyman (1951), Jagersten (1955), Hadzi (1963) など多くの動物学者が,ヒラムシはプラヌラに似た祖先株から進化し,アコエラは扁形動物の中で最も原始的なグループだと考えている(図14.27A)。
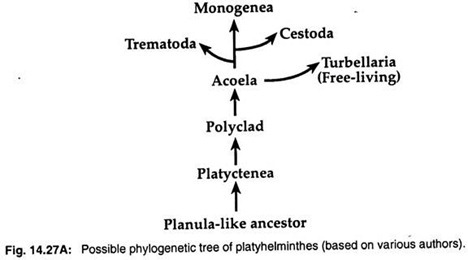
分子データとクラディスティック分析により、3つの寄生グループの起源は単系統であると示唆されている。 寄生する3つのクラス-Trematoda, Monogenea, Cestodaは自由生活するターベラーから進化したものである。
3.ここでも腸腔説は、両生類は基本的にすべて腸腔動物であり、無孔性扁形生物は腸腔動物の祖先から二次的に腔が失われて進化してきたとする。 この説は、腔腸動物から無腔動物への変化、両側対称から放射状対称への変化など、説明が難しい面があるため、動物学者には受け入れがたい。