免疫系
免疫系は組織、細胞、分子からなり、その主な機能は病原体と変質した細胞を検出、反応、排除することである。
自然免疫系と適応免疫系
免疫系には、1)自然免疫系、2)適応免疫系という2つの主要な構成要素があります。 自然免疫系は防御の第一線として働き、生殖細胞系列にコード化されたパターン認識受容体を介して病原体を検出し、記憶を形成せず、迅速に(数分から数時間以内に)反応する。 適応免疫系は、ゆっくりと(数日かけて)反応し、発生過程で何度も遺伝子再編成された抗原特異的受容体を使用し、免疫学的記憶を形成する-最初の攻撃から「学び」、将来同様の有害物質と戦うための準備をする。 どちらのタイプの免疫反応も、細胞外液に含まれる高分子によって、あるいは特定の免疫細胞の活性化によって引き起こされることがある。 これらの反応は、それぞれ体液性免疫、細胞媒介性免疫と呼ばれている。
細胞媒介性免疫応答の傘の下では、免疫細胞は、その応答が自然なものか適応的なものかに基づいて層別化されることがほとんどである。 自然免疫系は主に好中球、マクロファージ、ナチュラルキラー細胞、好塩基球などの食細胞からなり、危険な侵入生物を標的にする。 適応免疫系の細胞は、T細胞とB細胞である。 ナチュラルキラー(NK)T細胞は、自然免疫系と適応免疫系の両方の特徴を備えている。 さらに、免疫系に特異的でない他のほとんどの種類の細胞は、RIG-I、STING、NLRファミリーのメンバーのような細胞質レセプターとシグナル伝達およびエフェクター分子の形で、内在する自然免疫機能を保持することができる。 さらに検討するように、両系統の細胞は、造血幹細胞の異なる骨髄系とリンパ系の系統から生じる。
免疫系の構成要素
成熟した免疫細胞は、造血幹細胞として知られるものから分化します。 造血幹細胞は、骨髄、末梢血、胎盤に存在する多能性未分化前駆細胞である。 これらの造血幹細胞は、一般的な骨髄系前駆細胞や一般的なリンパ系前駆細胞に分化することができる。
リンパ系細胞
一般的なリンパ系前駆細胞は、細胞表面の受容体発現によって区別される4つの主要なリンパ球集団に分化することができる。 T細胞、B細胞、ナチュラルキラー(NK)細胞、およびNK-T細胞である。
T 細胞
T細胞は細胞媒介性免疫において重要な役割を担っている。 これらの細胞はCD3 T細胞シグナル伝達鎖の発現により同定される。
T細胞は、もともと骨髄の造血幹細胞から発生したリンパ系前駆細胞から発生します。 リンパ系前駆細胞はT細胞になることを決意すると、骨髄から胸腺に移動する(これがT細胞という名前の由来である)。 胸腺は、T細胞が細胞表面に発現する特異的な受容体によって定義される様々なサブタイプに成長するための適切な微小環境を提供する。 初期の成熟T細胞、すなわち胸腺細胞は、CD4とCD8の両方の受容体を持たないため、ダブルネガティブ(DN)細胞と呼ばれる。 DN細胞はその後、αおよびβT細胞受容体(TCR)をコードする遺伝子に遺伝子再配列や突然変異を起こし、最終的に固有の抗原に親和性を持つT細胞受容体となる系譜をたどることになる。
胸腺細胞は次に、細胞表面のMHCを介してペプチドとして提示される宿主抗原ではなく、「外来」抗原を認識するように、正と負の両方向から選別されます。 両方のテストに合格した細胞だけが、CD4とCD8を発現するようになり、二重陽性細胞となる。 成熟が終わると、T細胞はCD4かCD8のどちらかを発現するようになり(もう一方の発現は失われる)、単体陽性細胞となる。 これらの成熟したCD4またはCD8陽性T細胞は、その後血流中に放出されるが、最初はナイーブな状態、つまりまだ特定のT細胞サブタイプにコミットしていない状態である。
成熟中の胸腺細胞が自己のMHCペプチドに対して中程度の親和性を示す場合、有害な「自己」抗原を認識する能力を持つ制御性T細胞(Treg)になるよう選択されることがある。 血液中や末梢組織中のT細胞も、局所的にTregに誘導することができる。 Tregの主な役割は、自己抗原に対する寛容性を維持し、T-エフェクター細胞の機能と増殖を制限することである。
血液や末梢組織に到着したナイーブT細胞は、Tヘルパー細胞になるように誘導することができる。 マクロファージ、樹状細胞、B細胞などの抗原提示細胞(APC)がT細胞に抗原を示し、その受容体が親和性を持っていれば、T細胞は抗原を認識することができる。 通常、CD4+ T細胞は、クラスII MHCタンパク質に結合したペプチド抗原を認識するのに適している。 ペプチド-MHC複合体が認識されると、T細胞は一連の内部経路を活性化し、抗原認識を確認できるようにする。 このとき初めてTヘルパー細胞は増殖し、異なる細菌や寄生虫などの有害な抗原に特異的な利用可能な細胞のプールが拡大する。
Tヘルパー細胞が抗原を認識して活性化すると、さまざまなサイトカインを放出します。これは、他の免疫細胞に、有害な分子にどのように適切に反応するかのシグナルを送る分子です。 ヘルパーT細胞には、Th1、Th2、Th17といったいくつかのサブセットがある。 これらのサブセットは、病原体の種類に応じて免疫反応を調整するのに役立つ、異なるサイトカインを産生・分泌する。 一方、Th2細胞はIL-4、IL-5、IL-9、IL-10、IL-13を分泌し、好塩基球、好酸球、その他の細胞が感染症を攻撃するよう誘導する。 サイトカインの具体的な量と種類によって、感染症や有害な細胞に対する適切な反応が決定される。 T細胞は、細胞傷害性T細胞(またはナチュラルキラーT細胞)となって、自己の細胞に反応することもできる。 通常、これらはCD8+細胞であり、ウイルスに感染した細胞や腫瘍細胞を認識する。 ナチュラルキラーT細胞(NKT)については、後のセクションで詳しく説明する。 これらおよび他の種類のT細胞は、有害な物質を認識し、それを破壊するために他の免疫細胞を勧誘するとともに、同じ種類の侮辱に対して持続的な免疫応答を構築するために、さまざまな任務またはエフェクター機能をばらばらに実行する。
T細胞受容体シグナル相互作用経路については、ここで詳しくレビューしています。
CD3ε (D7A6E™) XP® Rabbit mAb #85061: IHC analysis of paraffin-embedded human breast carcinoma using #85061を使用した解析。
CD3ε (D7A6E™) XP® Rabbit mAb #85061: IHC分析(パラフィン包埋ヒト乳癌) #85061 を使用した分析。
CD4 (D7D2Z) Rabbit mAb #25229: IHC analysis of paraffin-embeded 4T1 metastatic tumor in mouse lung using #25229.
CD4 (D7D2Z) Rabbit mAb #25229: IHC analysis of paraffin-embedded 4T1 metastastatic tumor in mouse lung using #25229.
CD8α (D4W2Z) XP® Rabbit mAb (Mouse Specific) #98941を使用したマウス肺転移腫瘍のパラフィン包埋IHC解析。 98941 を用いたマウス肺のパラフィン包埋 IHC 解析。
CD8α (D4W2Z) XP® Rabbit mAb (Mouse Specific) #98941 を用いたマウス肺のパラフィン包埋 IHC 解析。 98941 を用いたマウス肺のパラフィン包埋 IHC 解析。
FoxP3 (D6O8R) Rabbit mAb #12653: IHC analysis of paraffin-embeded 4T1 metastatic tumor in mouse lung using #12653.
FoxP3 (D6O8R) Rabbit mAb #12653: IHC analysis of paraffin-embedded 4T1 metastastatic tumor in mouse lung using #12653.
B 細胞
B細胞は、血液中で抗体産生を仲介する重要な細胞である。 血液中に有害な物質が存在すると、B細胞上の特異的な受容体によって抗原として認識される。 そして、抗原を処理した後、T細胞の助けを借りて、B細胞は抗体を分泌する血漿細胞へと成熟する。 B細胞は、抗原の提示やサイトカイン分泌の役割も果たすことができる。 B細胞の発生と様々な機能の両方について説明する。
末梢血中に見出される白血球の15%はB細胞である。 B細胞は、2本の同一の重鎖と2本の同一の軽鎖からなる抗原結合タンパク質(抗体とも呼ばれる)である免疫グロブリン(Ig)を産生する。 これらの免疫グロブリンは、抗体分子ごとにアミノ酸配列が異なる特異的な抗原結合部位を有している。 これらの部位は、各抗体が異なる抗原を認識するための特異性を付与する。
上述のように、B細胞は骨髄に存在する造血細胞から分化します。 骨髄の中で、表面のIg受容体(抗体)が遺伝子再配列の過程を経て組み立てられているのである。 B細胞は、Igを構成する重鎖と軽鎖の異なる部分をコードする、多数の染色体上の異なる遺伝子の体細胞結合を利用している。 B細胞は、骨髄間質細胞によるIL-7の産生によって分化へと向かう。 これらの発達中のB細胞は、細胞表面の受容体発現の差によって媒介される重鎖および軽鎖の連続的な再配列のプログラムに従う。 例えば、特定のプレB細胞受容体の発現は、成熟B細胞が単一の抗原を特異的に認識できる特定のIg表面受容体を発現できるように、無関係な染色体上の特定の遺伝子の再配列を阻止する。 この過程は対立遺伝子排除と呼ばれている。 Ig重鎖と軽鎖が正しく組み立てられないと、受容体編集のための二次的な再配列が引き起こされることがある。
Ig受容体の細胞表面での発現とは別に、B細胞は、抗原がIg受容体の細胞外部分に結合した後に細胞内シグナルを誘導できる膜貫通型タンパク質も含んでいる。 B細胞は、T細胞と同様の細胞内シグナル伝達カスケードを用いて、増殖や成熟につながる転写変化を誘導している。 具体的には、Ig受容体の細胞内ドメインはキナーゼのsrcファミリーによってリン酸化され、チロシンキナーゼSykとBLINKとして知られるリンカータンパク質によってホスホリパーゼCとグアニン交換因子に受け渡されるカスケードを開始し、最終的にはプロテインキナーゼCを活性化してカルシウムを動員し、Ras/Rac依存的方法でMAPキナーゼを活性化させることができる。
B細胞のユニークな特徴は、アイソタイプスイッチングを行う能力である。 これは特に、わずかに異なる重鎖を生成することができるDNA再配列のプロセスを指す。 さらに、重鎖エキソンの代替スプライシングは、膜貫通エキソンがスプライシングされた場合、Igが細胞表面に結合したままであるか、または分泌されるようになることを可能にすることができる。 選択的スプライシングとアイソタイプスイッチングは、T細胞由来のサイトカインによって誘導される。 さらに、T細胞はB細胞のIgの抗原結合部位に変化をもたらす体細胞変異を誘発することがある。 B細胞の自己修正機能は、これらの一見ランダムに見える突然変異が、抗原に対する親和性の喪失につながるかどうかを検出し、細胞死のプログラムを開始させることができる。 このように、T細胞とB細胞の両方を活性化する抗原は、アイソタイプスイッチングと体細胞突然変異を通じて、T細胞を介したB細胞の成熟を誘発するが、この両者はB細胞の記憶と高度に関連しているのである。
記憶反応は、このような抗原に対する迅速かつ広範な高親和性Ig産生によって特徴づけられる。 このような記憶応答の発現は、ワクチン接種の成功に不可欠である。 しかしながら、これらは自己免疫やアレルギー反応を増加させることもある。
抗原によるT細胞独立型のB細胞活性化も起こりうるが、この種の活性化は弱い免疫記憶と結びついている。 さらに、これらの抗原は、B細胞を活性化するために高分子でなければならない。
NB: サイトカイン、またはインターロイキン(IL)は、膜貫通型の細胞表面受容体に作用する分泌分子である。 結合すると、これらのサイトカインは、Jak-STATシグナルに依存する典型的な細胞内シグナル伝達経路を活性化する。
B細胞受容体シグナル伝達相互作用経路については、ここで詳しくレビューしている。
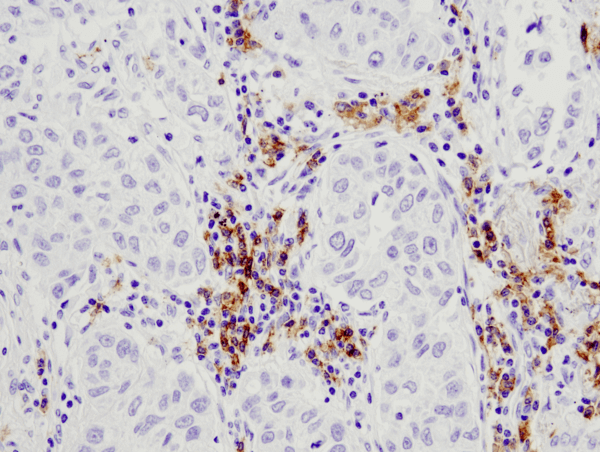
CD19 (D4V4B) XP® Rabbit mAb #90176: IHC analysis of paraffin-embedded human lung carcinoma using #90176.D4V4B XP Rabbit mAb #90176: IHC analysis of paraffin-embedded human lung carcinoma using #90176.
CD19 (D4V4B) XP® Rabbit mAb #90176: IHC analysis of paraffin-embedded human lung carcinoma using #90176.D4V4B XP mAb #90176: IHC analysis of paraffin-embedded human lung carcinoma using #90176.
NK 細胞
ナチュラルキラー(NK)細胞は、リンパ系から派生したもう一つの細胞タイプです。 B細胞と同様に、サイトカインと骨髄間質細胞に依存した方法で骨髄で発生する。 末梢血に占める割合は低く、抗原特異的な受容体を持たない。 その代わり、NK細胞は複雑な膜貫通型受容体のセットに依存している。 特に、クラスI HLA分子を認識する抑制性細胞表面受容体を持ち、そのため自己MHC分子によって抑制され、クラスI HLAの発現が低下した細胞のみを殺傷するのである。 したがって、CD8+ナチュラルキラーT(NK-T)細胞による死を回避するためにクラスI HLAの発現が低下したウイルス感染細胞や腫瘍細胞を標的にするという重要な役割を担っているのである。 NK細胞はまた、抗体、インターフェロン、サイトカイン依存的に活性化されるため、腫瘍との闘いにおいて大きな役割を果たすことができる。 NK細胞の作用機序は、細胞質から放出される顆粒酵素やパーフォリンを含む小顆粒に依存する。 これらのタンパク質は、細胞内に孔を開け、細胞内のタンパク質を分解し、アポトーシスを誘導することができる。 このように、NK細胞は細胞傷害性を持つと言われている。
NK-T細胞は、T細胞受容体の限られたレパートリーと組み合わせて、NK細胞が通常発現する特定の受容体およびマーカーを発現する、特別なサブタイプのT細胞である。 したがって、このユニークなサブタイプは、T細胞とNK細胞の両方の特性を共有している。 骨髄で発生するNK細胞とは異なり、NK-T細胞は胸腺で発生する。
骨髄系細胞
造血幹細胞の子孫である共通の骨髄系前駆細胞は、マクロファージ、樹状細胞、その他いくつかの自然免疫細胞や血液細胞を含むいくつかの異なるタイプの骨髄系細胞を生じます。
マクロファージ
マクロファージは、傷ついた細胞や異物などの抗原を認識し、オンデマンドで破壊するための引き金を引くことができます。 マクロファージはほとんどの組織に存在し、感染症や死にかけの細胞に対して必要な時に反応する。 認識された物質はマクロファージ内で貪食により破壊され、これがこの細胞の名前の由来となっている(ギリシャ語で「大きな食べる人」)。 マクロファージは、さまざまな場所に存在するとさまざまな形態をとり、貪食以外の機能も果たすことができる。
組織の損傷や病原体の感染に伴い、血液中の単球が患部組織に集められ、分化してマクロファージとなる。 組織の局在によって、肝臓のクッパー細胞、肺の肺胞マクロファージ、脳のミクログリアなど、さまざまな種類のマクロファージが存在する。 これらの異なるタイプのマクロファージは、いずれも単球に由来するが、その機能を常在組織に特化させたものである。 一般的な貪食機能のほとんどは、組織常駐型のマクロファージが担っている。 マクロファージは死細胞や異物を貪食する以外に、サイトカインを介して他の免疫細胞にシグナルを送ることもできる。 また、マクロファージは抗原提示という重要な機能をある程度担っており、T細胞と連携して適応免疫を支えている。 さらに、マクロファージはIL-12などのサイトカインを分泌して局所的な免疫反応を担うこともあれば、IL-10を大量に分泌して組織の修復に関わることもある。 このように、「大食漢」は、免疫系において、主な仕事である貪食に加えて、さまざまな役割を担っているのである。
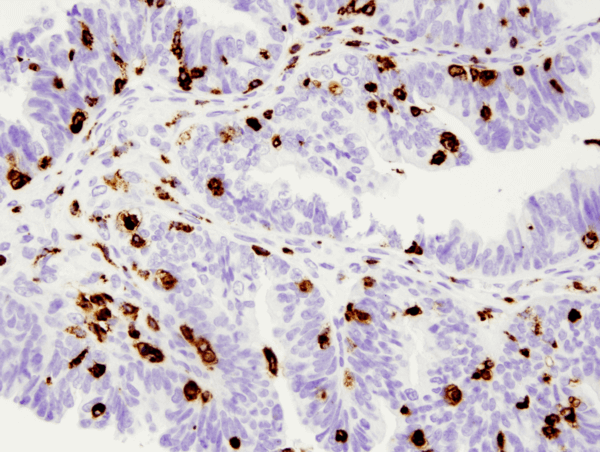
CD68 (D4B9C) XP® Rabbit mAb #76437: IHC analysis of paraffin-embedded human serous papillary carcinoma of the ovary using #76437.は、CD68 (D4B9C) XP® Rabbit mAb #76437 を用いて作成されました。
CD68 (D4B9C) XP® Rabbit mAb #76437: IHC analysis of paraffin-embedded human serous papillary carcinoma of the ovary using #76437.による卵巣の漿液性乳頭癌のIHC分析。
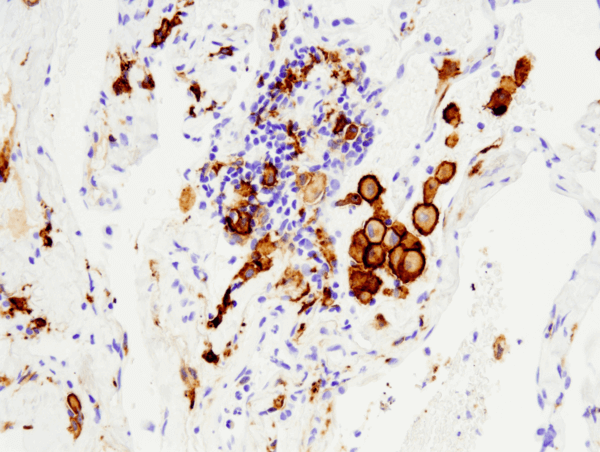
CD163 (D6U1J) Rabbit mAb #93498: Leica® Bond™ Rxで行った#93498を用いたパラフィン包埋ヒト非小細胞肺癌のIHC解析です。
CD163 (D6U1J) Rabbit mAb #93498: IHC analysis of paraffin-embedded human non-small cell lung carcinoma using #93498 on the Leica® Bond™ Rx.Lab.(Leica® Bond® Rx.Lab.
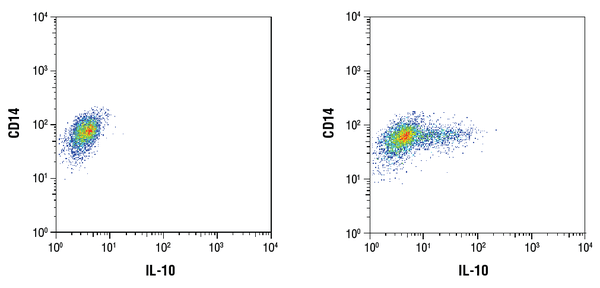
IL-10 (D13A11) XP® Rabbit mAb #12163: 精製したCD14+ヒト末梢血単核細胞を未処理(左)またはLPSで処理(100ng/mL、16時間;右)し、CD14抗体と#12163を用いたフローサイトメトリー解析。 LPS刺激1時間後にBrefeldin A #9972 (300ng/mL) を未処理細胞および処理細胞に添加した。 二次抗体として Anti-rabbit IgG (H+L), F(ab’)2 Fragment (Alexa Fluor® 647 Conjugate) #4414 を使用した。
IL-10 (D13A11) XP® Rabbit mAb #12163: 精製したCD14+ヒト末梢血単核細胞を未処理(左)またはLPSで処理(100ng/mL、16時間;右)し、CD14抗体と#12163を用いたフローサイトメトリー解析。 LPS刺激1時間後にBrefeldin A #9972 (300ng/mL) を未処理細胞および処理細胞に添加した。 二次抗体として Anti-rabbit IgG (H+L), F(ab’)2 Fragment (Alexa Fluor® 647 Conjugate) #4414 を使用した。
Dendritic Cells
マクロファージと同様に、樹状細胞は細胞や異物を取り込む能力を持ちますが、除去のために消化するのではなく、物質を加工して抗原としてT細胞へ提示するのが樹状細胞です。 このように、樹状細胞は抗原提示細胞(アクセサリー細胞)として、自然免疫系と適応免疫系の間で病原体に関する情報を中継する役割を担っているのである。 樹状細胞は、その明確な細胞突起が手を伸ばし、有害物質を掴んで消化することから、その名が付けられた。
造血器系の骨髄前駆細胞から発生した未熟な樹状細胞は、最初は「学習」段階を経て、異物がないかどうかを局所的にサンプリングし、パターン認識受容体(これらの受容体の共通セットの1つはtoll様受容体)を介して認識応答を開始します。 これらの未熟な樹状細胞は、鼻の奥、胃、肺、さらには皮膚など、外部環境にアクセスできるさまざまな場所に存在することができます。 抗原を認識すると、樹状細胞はリンパ節に移動し、そこでT細胞に抗原を認識させ、それに応じて反応するよう訓練する。 樹状細胞は、メモリーT細胞とナイーブT細胞の両方を活性化する唯一の抗原提示細胞である。 Toll様受容体は、マクロファージや樹状細胞に存在し、外来病原体の認識と応答を媒介する、よく研究されている受容体の一種である。 樹状細胞は、T細胞と直接相互作用する以外にも、IL-12などのサイトカインを介してシグナルを送ることができ、有害物質に対するその後の攻撃のために免疫系を準備するよう作用する。 他の種類の樹状細胞は、過剰な廃棄物を一掃するために、より多くのマクロファージを呼び寄せるシグナルとなるサイトカインを分泌することもある。 このように、樹状細胞は抗原提示の主要な細胞メディエーターとして、自然免疫系と適応免疫系の間のクロストークに不可欠な役割を担っているのである。
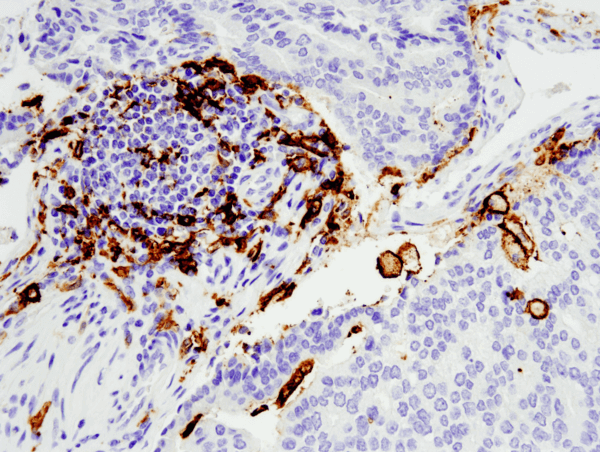
CD11c (D3V1E) XP® Rabbit mAb #45581: Leica® Bond™ Rxで実施した#455812を用いたパラフィン包埋ヒト前立腺腺癌のIHC解析。
CD11c (D3V1E) XP® Rabbit mAb #45581: 455812 を用いたパラフィン包埋ヒト前立腺腺癌の IHC 解析(Leica® Bond™ Rx で実施)。
その他の自然免疫系細胞とシグナル伝達
自然免疫系のその他の細胞には、創傷治癒に働き、病原体に対する防御を助けるマスト細胞が含まれます。 病原体によって引き起こされると、マスト細胞はサイトカインを分泌するとともに、ヒスタミンやヘパリンを含む顆粒を分泌し、マクロファージの動員や血管の拡張など、さらなる免疫反応を媒介することができる。 好中球、好酸球、好塩基球は、細胞質内に特徴的な顆粒を持つため、総称して顆粒球と呼ばれる。 好中球の場合、顆粒は真菌や細菌などの病原体を標的とした毒性化合物を放出する働きをする。
Immunology in Health and Disease
Cancer Immunology
免疫細胞を利用してがんと戦うことは、臨床での成功により、非常にホットな研究テーマになっています。 現在の免疫療法の焦点は、通常T細胞の活性化と反応を妨げるPD-1などのシグナル伝達チェックポイントタンパク質を標的とすることである。 養子縁組細胞療法は、患者さんから採取したT細胞を免疫編集する、もう一つのエキサイティングなアプローチです。 このT細胞は、患者さんのがん細胞に特異的なキメラ抗原受容体を発現するように遺伝子操作されます。 T細胞はその後、患者に再注入され、通常のT細胞の機能に従って、がん細胞を探し出し、殺すようにプログラムされます。 このように、免疫系を操作して患者の有害ながん細胞を攻撃するアプローチは、がん治療のための画期的な治療法であるといえます。