Résultats d’apprentissage
- Identifier les causes historiques et potentielles des taux d’extinction élevés
Le nombre d’espèces sur la planète, ou dans n’importe quelle zone géographique, est le résultat d’un équilibre de deux processus évolutifs qui sont continuellement en cours : la spéciation et l’extinction. Tous deux sont des processus naturels de « naissance » et de « mort » de la macroévolution. Lorsque le taux de spéciation commence à dépasser le taux d’extinction, le nombre d’espèces augmente ; de même, le nombre d’espèces diminue lorsque le taux d’extinction commence à dépasser le taux de spéciation. Tout au long de l’histoire de la Terre, ces deux processus ont fluctué – entraînant parfois des changements spectaculaires dans le nombre d’espèces sur Terre, comme le reflète le registre fossile (figure 1).
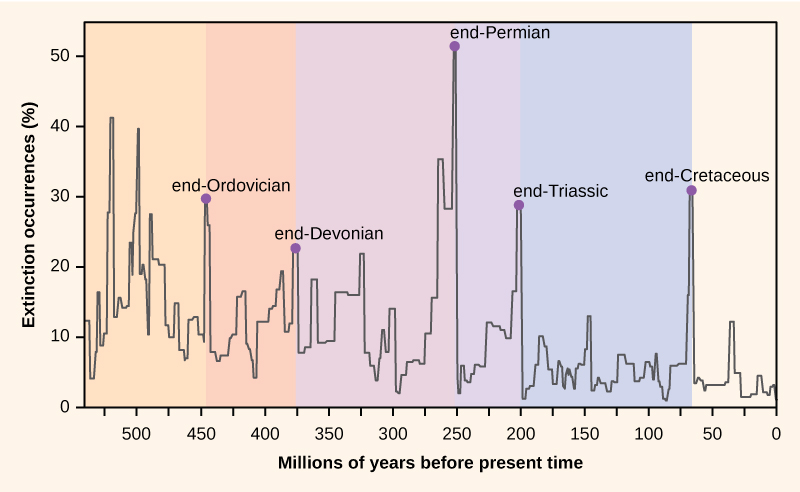
Figure 1. Le pourcentage d’occurrences d’extinction tel que reflété dans le registre fossile a fluctué tout au long de l’histoire de la Terre. Des pertes soudaines et dramatiques de biodiversité, appelées extinctions de masse, se sont produites cinq fois.
Les paléontologues ont identifié cinq strates dans le registre fossile qui semblent présenter des pertes soudaines et dramatiques (plus de la moitié de toutes les espèces existantes disparaissant du registre fossile) de biodiversité. On les appelle des extinctions massives. Il existe de nombreux événements d’extinction moins importants, mais tout aussi spectaculaires, mais les cinq extinctions massives sont celles qui ont suscité le plus de recherches. On peut avancer que les cinq extinctions massives ne sont que les cinq événements les plus extrêmes d’une série continue de grands événements d’extinction tout au long du Phanérozoïque (depuis 542 millions d’années). Dans la plupart des cas, les causes hypothétiques sont encore controversées ; cependant, l’événement le plus récent semble clair.
Extinctions de masse enregistrées
Les archives fossiles des extinctions de masse ont servi de base pour définir les périodes de l’histoire géologique, elles se produisent donc généralement au point de transition entre les périodes géologiques. La transition dans les fossiles d’une période à l’autre reflète la perte dramatique d’espèces et l’origine graduelle de nouvelles espèces. Ces transitions sont visibles dans les strates rocheuses. Le tableau 1 fournit des données sur les cinq extinctions de masse.
| Tableau 1. Extinctions de masse | ||
|---|---|---|
| Période géologique | Nom de l’extinction de masse | Moment (il y a des millions d’années) |
| Ordovicien-Silurien | fin.Ordovicien O-S | 450-440 |
| Dévonien tardif | suite-Dévonien | 375-360 |
| Permien-Trias | en-Permien | 251 |
| Trias-Jurassique | en-…Trias | 205 |
| Crétacé-Paléogène | end-Crétacé K-Pg (K-T) | 65.5 |
L’événement d’extinction Ordovicien-Silurien est la première extinction de masse enregistrée et la deuxième plus grande. Au cours de cette période, environ 85 % des espèces marines (peu d’espèces vivaient en dehors des océans) se sont éteintes. La principale hypothèse quant à sa cause est une période de glaciation puis de réchauffement. Le phénomène d’extinction se compose en fait de deux événements d’extinction séparés d’environ 1 million d’années. Le premier événement a été causé par un refroidissement, et le second événement est dû au réchauffement qui a suivi. Les changements climatiques ont affecté les températures et le niveau des mers. Certains chercheurs ont suggéré qu’un sursaut gamma, provoqué par une supernova proche, était une cause possible de l’extinction de l’Ordovicien et du Silurien. Le sursaut gamma aurait enlevé la couche d’ozone protectrice de la Terre, permettant aux rayons ultraviolets intenses du soleil d’atteindre la surface de la Terre, et pourrait expliquer les changements climatiques observés à l’époque. Cette hypothèse est très spéculative, et les influences extraterrestres sur l’histoire de la Terre constituent une ligne de recherche active. La récupération de la biodiversité après l’extinction de masse a pris de 5 à 20 millions d’années, selon les endroits.
L’extinction de la fin du Dévonien a pu se produire sur une période relativement longue. Elle semble avoir surtout affecté les espèces marines et pas tellement les plantes ou les animaux habitant les habitats terrestres. Les causes de cette extinction sont mal comprises.
L’extinction de la fin du Permien a été la plus importante de l’histoire de la vie. En effet, on pourrait soutenir que la Terre est devenue presque dépourvue de vie pendant cet événement d’extinction. On estime que 96 % de toutes les espèces marines et 70 % de toutes les espèces terrestres ont disparu. C’est à cette époque, par exemple, que les trilobites, un groupe qui avait survécu à l’extinction de l’Ordovicien-Silurien, se sont éteints. Les causes de cette extinction massive ne sont pas claires, mais le principal suspect est une activité volcanique étendue et généralisée qui a conduit à un réchauffement global incontrôlé. Les océans sont devenus largement anoxiques, étouffant la vie marine. La diversité des tétrapodes terrestres a mis 30 millions d’années à se rétablir après l’extinction de la fin du Permien. L’extinction du Permien a radicalement modifié la composition de la biodiversité de la Terre et le cours de l’évolution.
Les causes de l’événement d’extinction du Trias-Jurassique ne sont pas claires, et les chercheurs avancent des hypothèses incluant le changement climatique, l’impact d’astéroïdes et les éruptions volcaniques. L’événement d’extinction s’est produit juste avant la rupture du supercontinent Pangée, bien que des études récentes suggèrent que les extinctions ont pu se produire plus progressivement tout au long du Trias.
Les causes de l’événement d’extinction de la fin du Crétacé sont celles qui sont les mieux comprises. C’est au cours de cet événement d’extinction, il y a environ 65 millions d’années, que la majorité des dinosaures, le groupe de vertébrés dominant pendant des millions d’années, a disparu de la planète (à l’exception d’un clade de théropodes qui a donné naissance aux oiseaux).
La cause de cette extinction est maintenant comprise comme étant le résultat d’un impact cataclysmique d’une grande météorite, ou astéroïde, au large de la côte de ce qui est maintenant la péninsule du Yucatán. Cette hypothèse, proposée pour la première fois en 1980, était une explication radicale fondée sur un pic brutal des niveaux d’iridium (qui pénètre dans notre atmosphère à partir de météores à un rythme assez constant mais qui est par ailleurs absent de la surface de la Terre) dans la strate rocheuse qui marque la limite entre les périodes du Crétacé et du Paléogène (figure 2). Cette limite a marqué la disparition des dinosaures dans les fossiles ainsi que de nombreux autres taxons. Les chercheurs qui ont découvert le pic d’iridium l’ont interprété comme un afflux rapide d’iridium de l’espace vers l’atmosphère (sous la forme d’un gros astéroïde) plutôt que comme un ralentissement du dépôt de sédiments au cours de cette période. C’était une explication radicale, mais le rapport d’un cratère d’impact d’âge et de taille appropriés en 1991 a rendu l’hypothèse plus crédible. Aujourd’hui, de nombreuses preuves géologiques viennent étayer cette théorie. Les temps de récupération de la biodiversité après l’extinction de la fin du Crétacé sont plus courts, en temps géologique, que pour l’extinction de la fin du Permien, de l’ordre de 10 millions d’années.
Une autre possibilité, qui coïncide peut-être avec l’impact de l’astéroïde du Yucatan, est un volcanisme étendu qui a commencé à se former il y a environ 66 millions d’années, à peu près en même temps que l’impact de l’astéroïde du Yucatan, à la fin du Crétacé. Les coulées de lave ont recouvert plus de 50 % de ce qui est aujourd’hui l’Inde. Le dégagement de gaz volcaniques, en particulier de dioxyde de soufre, pendant la formation des pièges a contribué au changement climatique, qui pourrait avoir induit l’extinction massive.
Question pratique

Figure 2. Bande d’iridium (crédit : USGS)
En 1980, Luis et Walter Alvarez, Frank Asaro et Helen Michels ont découvert, à travers le monde, un pic de concentration d’iridium au sein de la couche sédimentaire à la limite K-Pg. Ces chercheurs ont émis l’hypothèse que ce pic d’iridium avait été provoqué par l’impact d’un astéroïde ayant entraîné l’extinction de masse K-Pg. Dans la figure 2, la couche d’iridium est la bande claire.
Les scientifiques ont mesuré l’abondance relative des spores de fougère au-dessus et au-dessous de la limite K-Pg dans cet échantillon de roche. Lequel des énoncés suivants représente le plus probablement leurs résultats ?
- Une abondance de spores de fougère de plusieurs espèces a été trouvée sous la limite K-Pg, mais aucune n’a été trouvée au-dessus.
- Une abondance de spores de fougère de plusieurs espèces a été trouvée au-dessus de la limite K-Pg, mais aucune n’a été trouvée au-dessous.
- Une abondance de spores de fougères a été trouvée à la fois au-dessus et au-dessous de la limite K-Pg, mais une seule espèce a été trouvée au-dessous de la limite, et de nombreuses espèces ont été trouvées au-dessus de la limite.
- De nombreuses espèces de spores de fougères ont été trouvées à la fois au-dessus et au-dessous de la limite, mais le nombre total de spores était plus important au-dessous de la limite.
L’extinction pléistocène
L’extinction pléistocène est l’une des plus petites extinctions, et une extinction récente. Il est bien connu que la mégafaune nord-américaine, et dans une certaine mesure eurasienne, – de grands animaux vertébrés – a disparu vers la fin de la dernière période de glaciation. L’extinction semble s’être produite au cours d’une période relativement restreinte, il y a 10 000 à 12 000 ans. En Amérique du Nord, les pertes ont été assez spectaculaires et ont concerné les mammouths laineux (avec une population existante jusqu’à il y a environ 4 000 ans, isolée sur l’île Wrangel, au Canada), le mastodonte, les castors géants, les paresseux géants, les chats à dents de sabre et le chameau d’Amérique du Nord, pour n’en citer que quelques-uns. Au début des années 1900, les scientifiques ont suggéré pour la première fois que la chasse excessive avait causé l’extinction rapide de ces grands animaux. Les recherches sur cette hypothèse se poursuivent aujourd’hui.
En général, la chronologie des extinctions du Pléistocène est en corrélation avec l’arrivée des paléo-humains, peut-être il y a 40 000 ans, et non avec les événements liés au changement climatique, qui est la principale hypothèse concurrente pour ces extinctions. Les extinctions ont commencé en Australie il y a environ 40 000 à 50 000 ans, juste après l’arrivée des humains dans la région : un lion marsupial, un wombat géant d’une tonne et plusieurs espèces de kangourous géants ont disparu. En Amérique du Nord, l’extinction de presque tous les grands mammifères a eu lieu il y a 10 000 à 12 000 ans. Il ne reste plus que les petits mammifères tels que les ours, les élans, les orignaux et les couguars. Enfin, sur de nombreuses îles océaniques éloignées, l’extinction de nombreuses espèces a coïncidé avec l’arrivée de l’homme. Toutes les îles ne comptaient pas de grands animaux, mais lorsqu’il y en avait, ils étaient souvent contraints à l’extinction. Madagascar a été colonisée il y a environ 2 000 ans et les grands mammifères qui y vivaient se sont éteints. L’Eurasie et l’Afrique ne présentent pas ce schéma, mais elles n’ont pas non plus connu une arrivée récente d’humains chasseurs-cueilleurs. L’homme est arrivé en Eurasie il y a des centaines de milliers d’années. Ce sujet reste un domaine de recherche et d’hypothèses actif. Il semble clair que même si le climat a joué un rôle, dans la plupart des cas, la chasse humaine a précipité les extinctions.
Extinctions de l’époque actuelle
La sixième extinction de masse, ou Holocène, semble avoir commencé plus tôt qu’on ne le pensait et est largement due aux activités perturbatrices de l’Homo sapiens moderne. Depuis le début de la période de l’Holocène, il existe de nombreuses extinctions récentes d’espèces individuelles qui sont enregistrées dans les écrits humains. La plupart d’entre elles coïncident avec l’expansion des colonies européennes depuis les années 1500.
L’un des exemples les plus anciens et les plus connus est celui du dodo. Cet étrange oiseau ressemblant à un pigeon vivait dans les forêts de l’île Maurice (une île de l’océan Indien) et s’est éteint vers 1662. Le dodo était chassé pour sa viande par les marins et était une proie facile car il s’approchait des gens sans crainte (le dodo n’avait pas évolué avec les humains). Les cochons, les rats et les chiens apportés sur l’île par les navires européens tuaient également les jeunes et les œufs de dodo.
La vache de mer de Steller s’est éteinte en 1768 ; elle était apparentée au lamantin et vivait probablement autrefois le long de la côte nord-ouest de l’Amérique du Nord. La vache de mer de Steller a été découverte par les Européens en 1741 et a fait l’objet d’une chasse excessive pour la viande et l’huile. La dernière vache de mer a été tuée en 1768. Cela représente seulement 27 ans entre le premier contact de la vache de mer avec les Européens et l’extinction de l’espèce !
Depuis 1900, diverses espèces se sont éteintes, dont les suivantes :
- En 1914, le dernier pigeon voyageur vivant est mort dans un zoo de Cincinnati, en Ohio. Cette espèce avait autrefois obscurci le ciel de l’Amérique du Nord lors de ses migrations, mais elle a été chassée à outrance et a souffert de la perte d’habitat résultant du défrichement des forêts pour les terres agricoles.
- La perruche de Caroline, autrefois commune dans l’est des États-Unis, s’est éteinte en 1918. Elle a souffert de la perte de son habitat et a été chassée pour l’empêcher de manger les fruits des vergers. (La perruche mangeait les fruits des vergers parce que ses aliments indigènes ont été détruits pour faire place aux terres agricoles)
- Le lion de mer japonais, qui habitait une large zone autour du Japon et de la côte de la Corée, s’est éteint dans les années 1950 à cause des pêcheurs.
- Le phoque moine des Caraïbes était réparti dans toute la mer des Caraïbes mais a été conduit à l’extinction par la chasse en 1952.
Ce ne sont là que quelques-unes des extinctions enregistrées au cours des 500 dernières années. L’Union internationale pour la conservation de la nature (UICN) tient une liste des espèces éteintes et menacées, appelée liste rouge. Cette liste n’est pas complète, mais elle décrit 380 espèces éteintes de vertébrés après 1500 après JC, dont 86 ont été chassées par la surchasse ou la surpêche.
Estimations des taux d’extinction en temps présent
Les estimations des taux d’extinction sont entravées par le fait que la plupart des extinctions se produisent probablement sans observation. L’extinction d’un oiseau ou d’un mammifère est susceptible d’être remarquée par les humains, surtout s’il a été chassé ou utilisé d’une autre manière. Mais il existe de nombreux organismes qui présentent moins d’intérêt pour l’homme (pas nécessairement moins de valeur) et beaucoup qui ne sont pas décrits.
Le taux d’extinction de fond est estimé à environ un par million d’espèces par an (E/MSY). Par exemple, en supposant qu’il y a environ dix millions d’espèces en existence, on s’attend à ce que dix espèces s’éteignent chaque année (chaque année représente dix millions d’espèces par an).
Une estimation contemporaine du taux d’extinction utilise les extinctions dans les archives écrites depuis l’an 1500. Pour les oiseaux seulement, cette méthode donne une estimation de 26 E/MSY. Cependant, cette valeur peut être une sous-estimation pour trois raisons. Tout d’abord, de nombreuses espèces n’auraient été décrites que bien plus tard dans la période considérée, de sorte que leur disparition serait passée inaperçue. Deuxièmement, le nombre d’espèces de vertébrés récemment disparues augmente car les espèces disparues sont maintenant décrites à partir de restes de squelettes. Enfin, certaines espèces sont probablement déjà éteintes, même si les défenseurs de l’environnement hésitent à les désigner comme telles. Si l’on tient compte de ces facteurs, le taux d’extinction estimé se rapproche de 100 E/MSY. Le taux prévu d’ici la fin du siècle est de 1500 E/MSY.

Figure 3. Des études ont montré que le nombre d’espèces présentes augmente avec la taille de l’habitat. (crédit : modification des travaux d’Adam B. Smith)
Une deuxième approche pour estimer les taux d’extinction actuels consiste à corréler la perte d’espèces avec la perte d’habitat en mesurant la perte de surface forestière et en comprenant les relations espèces-surface. La relation espèce-zone est le taux auquel de nouvelles espèces sont observées lorsque la zone étudiée est augmentée. Des études ont montré que le nombre d’espèces présentes augmente avec la taille de l’île. Ce phénomène s’est également avéré vrai dans d’autres habitats de type insulaire, tels que les tepuis du Venezuela, qui sont entourés de forêt tropicale. En inversant cette relation, si la superficie de l’habitat est réduite, le nombre d’espèces qui y vivent diminuera également. Les estimations des taux d’extinction basées sur la perte d’habitat et les relations entre les espèces et la superficie ont suggéré qu’avec une perte d’habitat d’environ 90 %, 50 % des espèces disparaîtraient. Les estimations de la superficie des espèces ont conduit à des calculs du taux d’extinction des espèces d’environ 1000 E/MSY et plus. En général, les observations réelles ne montrent pas cette quantité de perte et il a été suggéré qu’il y a un retard dans l’extinction. Des travaux récents ont également remis en question l’applicabilité de la relation entre l’espèce et l’aire pour estimer la perte d’espèces. Ces travaux soutiennent que la relation espèce-espace conduit à une surestimation des taux d’extinction. Une meilleure relation à utiliser pourrait être la relation entre les espèces endémiques et la superficie. L’utilisation de cette méthode ramènerait les estimations à environ 500 E/MSY au cours du siècle à venir. Notez que cette valeur est encore 500 fois le taux de fond.
Essayez-le
Contribuez !
Améliorer cette pageEn savoir plus
.