ADVERTISEMENTS:
Tässä artikkelissa keskustelemme Platyhelminthes:- 1. Platyhelminthesin elinympäristö ja elinympäristö 2. Platyhelminthes. Platyhelminthesin rakenne 3. Tartuntaelimet 4. Runkoseinä 5. Ruoansulatusjärjestelmä 6. Eritysjärjestelmä 7. Hengityselimet 8. Hermosto 9. Lisääntymisjärjestelmä 10. Kehitys 11. Fylogeneettiset näkökohdat.
- Platyhelmintien elinympäristö ja elinympäristö:
- Platyhelmintien rakenne:
- Platyhelminttisten tarttumis- tai imusolmukkeet:
- Platyhelmintien ruumiinseinä:
- Platyhelmintien ruoansulatuselimistö:
- Platyhelmintien ulosteperäinen järjestelmä:
- Platyhelmintien hengitys- ja verenkiertojärjestelmät:
- Platyhelmintien hermosto:
- Platyhelmintien lisääntymisjärjestelmä:
- Platyhelmintien kehitys:
- Phylogenetic Considerations of Platyhelminthes:
Platyhelmintien elinympäristö ja elinympäristö:
Platyhelminteet ovat enimmäkseen ekto- tai endoparasiittisia ja vain harvat ovat vapaana eläviä. Vapaasti elävät kuuluvat Turbellaria-luokkaan ja elävät makeassa vedessä, lammissa, järvissä, puroissa ja lähteissä. Joitakin niistä tavataan trooppisten ja subtrooppisten alueiden rannoilla. Trematoda- ja cestoda-lajit ovat totaalisia loisia. Aikuisvaiheessa ne loisevat selkärankaisia ja toukkavaiheessa ne esiintyvät selkärangattomien eläinten loisina.
Platyhelmintien rakenne:
ALKUPERÄISET:
Suvun jäsenet ovat yleensä pitkänomaisia. Polycladit ovat leveälehtisiä, kun taas heisimadot ovat litteitä ja nauhamaisia. Rungon ääriviivat ovat yleensä yksinkertaiset, mutta joidenkin trematodien ääriviivat ovat omituiset. Heisimadon rungot koostuvat useista neliömäisistä tai suorakaiteen muotoisista segmenteistä, joita kutsutaan proglottideiksi.
Anteriorinen proglottidi on pienin ja takimmainen proglottidi on suurin. Se tarkoittaa, että proglottidin koko kasvaa anteroposterioriseen suuntaan. Proglottidien läsnäolo antaa heisimadoille segmentoituneisuuden. Procerodes lobatassa esiintyy jonkinlaista pseudometamerismiä, jossa jotkin sisäelimet toistuvat.
Anteriorinen ja posteriorinen pää sekä dorsaalinen ja ventraalinen pinta ovat helposti tunnistettavissa. Usein etupää erottuu muusta kehosta ”pään” ja sitä seuraavan supistuneen ”kaulan” avulla.
Joissakin muodoissa ei ole selvää päätä, mutta etupää on havaittavissa aistinelinten avulla tai siitä, että sen liike suuntautuu eteenpäin liikkumisen aikana. Ventraalipinnalla on suu ja sukupuoliaukot, jos niitä on.
LÄHTEET:
Platyhelmintien koot vaihtelevat mikroskooppisen pienistä äärimmäisen pitkulaisiin, jopa 10-15 metriä pitkiin muotoihin (heisimadot). Suurin osa jäsenistä on kooltaan pieniä tai keskikokoisia. Yleisesti ottaen platyhelminteet ovat värittömiä tai valkoisia, Vapaasti elävät muodot ovat väriltään valkoisia, ruskeita, harmaita tai mustia. Joillakin polykladeilla ja maaplanarioilla on kirkkaita värejä, jotka ovat järjestäytyneet kuvioiksi.
Platyhelminttisten tarttumis- tai imusolmukkeet:
Platyhelminteillä on erilaisia tarttumis- ja kiinnittymiselimiä. Acetabulum eli imuelin ”imijän” muodossa on hyvin yleinen aikuisilla litteillä matoilla. Liekoissa on kaksi imukappaletta vatsanpuoleisella sivulla. Toinen näistä imukappaleista sijaitsee rungon etupuolella, ja sen asento on enemmän tai vähemmän kiinteä.
Kun taas toinen, takimmaiseksi imijäksi kutsuttu imijä, ei ole vakioasennossa. Paramphistomumissa imijä on sijainniltaan takimmainen, Echinostomassa ja Fasciolassa sijainti on siirtynyt enemmän etupuolelle.
Heisimatoilla tarttumiselimet ovat urien tai kuppien muodossa, ja ne sijaitsevat kefaalisessa päässä. Usein näiden urien läheisyydessä tai eversiibelissä poskiontelossa on koukkuja, jotka auttavat kiinnittymisessä. Myös joillakin vapaana elävillä Planarioilla esiintyy imusolmukkeita.
Platyhelmintien ruumiinseinä:
Platyhelminteiltä puuttuu ulko- tai sisäluuranko, joten niiden ruumis on pehmeä. Epidermis on yksikerroksinen ja joissakin tapauksissa se on synkyylinen. Epidermis voi olla kokonaan tai osittain säikeinen. Epidermiksen alapuolinen kerros koostuu pyöreistä, pitkittäisistä ja vinoista kuiduista koostuvasta lihaksistosta. Lihassyyt ovat sileitä. Kaikki elinten väliset tilat on täytetty pakkautuvilla soluilla, joita kutsutaan parenkyymiksi.
Platyhelmintien kovia osia ovat koukut ja piikit. Monet aikaisemmat tutkijat katsoivat, että monogenealaisten, digenealaisten ja cestodien ruumista peittää ”kynsinauha”.
Valomikroskopiassa näissä muodoissa esiintyvä sovittamattomuus antaa valomikroskopiassa väärän vaikutelman kynsinauhasta. Elektronimikroskooppiset tutkimukset ovat kuitenkin paljastaneet, että kehon uloin kerros on epidermis. Tämän kerroksen solut sisältävät mikrokondrioita ja pysyvät jatkuvina alla olevien solujen kanssa.
Platyhelmintien ruoansulatuselimistö:
Ruuansulatusjärjestelmä puuttuu turkulaarijärjestyksessä Acoela ja cestodeissa. Turbellaria- ja Trematoda-heimon ruuansulatuselimistöä edustavat suu, nielu ja suoli, joka päättyy sokeasti, ja näin ollen ruoansulatuselinten kokonaisasetelma muistuttaa läheisesti muurahaiseläinten ja ctenoforien asetelmaa.
Suu sijaitsee alkukantaisessa tilassa suunnilleen keskellä vatsaviivaa, mutta monissa litteissä matoissa suu on siirtynyt etupuolelle keskivatsaviivaa pitkin.
HAVAINNOT:
Suu puuttuu endoparasiittisissa rabdokokkeissa. Nielu on luonteeltaan stomodaealinen ja vahva lihaksikas putki. Siinä on vaihtelua suvun sisällä. Suoli on muodoltaan hyvin vaihteleva. Se voi olla yksinkertainen pussi tai siinä voi olla monimutkaisia haarautumia ja alahaarautumia. Peräaukkoja on harvoin.
Platyhelmintien ulosteperäinen järjestelmä:
Platyhelmintien eritejärjestelmä koostuu pääasiassa pitkittäisistä vesiastioista tai pitkittäisistä eritekanavista, joissa on useita erikoistuneita eritesoluja, joita kutsutaan liekkisoluiksi tai liekkisipuleiksi, joita kutsutaan kollektiivisesti protonephridioiksi. Vesiastioiden sijoittelu rungon sisällä vaihtelee suuresti.
Kolmikotiloissa on kaksi pitkittäistä verisuonta (kanavaa), jotka avautuvat ulospäin lukuisista huokosista (kuva 14.28G). Rhabdocoaleissa on kaksi lateraalista tai yksi mediaaniastia (kuva 14.28E). Keskisuoni avautuu ulospäin yhdestä huokosesta, joka sijaitsee rungon takaosassa.
Muodoissa, joissa on kaksi verisuonta, aukot ovat joko ventraalisella puolella tai nielussa. Joissakin muodoissa molemmat verisuonet yhdistyvät takaosassa ja avautuvat ulospäin yhdestä keskiaukosta. Pääkanava on laajalti haaroittunut, ja nefriidiporeen lisäksi siinä on ylimääräisiä erittäviä huokosia (kuva 14.28F).
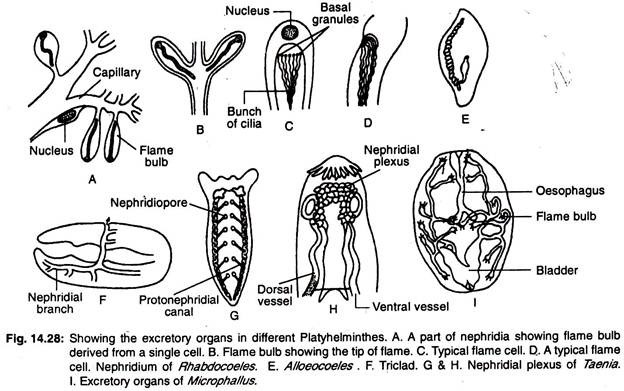
Yksisukuisilla trematodeilla on kaksi pitkittäistä ekskretorista kanavaa, ja molemmat kanavat avautuvat erikseen selkäpinnalle. Kanavat laajenevat muodostaen ulostepusseja päätepisteissään.
Muilla trematodeilla erityskanavat yhdistyvät posteriorisesti toisiinsa poikittaisella kanavalla ja avautuvat ulospäin yhdestä keskiaukosta. Poikittainen kanava dialisoituu ja muodostaa joissakin tapauksissa erittävän rakkulan. Kaksi pituussuuntaista kanavaa voi yhdistyä toisiinsa takaosassa, ja niissä voi olla yksi aukko.
Cestodeilla on neljä pitkittäistä erittymisverkkoa. Eritysverisuonet yhdistyvät scolexissa sijaitsevan poikittaisverisuonen avulla nephridinaalipleksiksi (kuva 14.28H) ja avautuvat viimeisessä proglottidissa sijaitsevaan supistuvaan eritysvesikkeliin.
Makean veden turbellariassa on lukuisia protonephridioita ja suuri määrä nephridioporeja, koska liiallisen veden tulo elimistöön estyy.
Platyhelmintien hengitys- ja verenkiertojärjestelmät:
HARJOITUKSET:
Nämä kaksi järjestelmää puuttuvat kokonaan heimosta. Joillakin trematodeilla on kuvattu lymfaattiseksi järjestelmäksi kutsutun putkijärjestelmän olemassaolo. Putkilla katsotaan olevan epävarmoja tehtäviä.
Platyhelmintien hermosto:
Päähermokeskus eli aivot sijaitsevat päässä parina aivokurkiaisena. Aivoista lähtee useita ganglionisia pitkittäishermosäikeitä. Näistä pitkittäisjuosteista pari tulee selvimmin esiin ja loput muuttuvat merkityksettömiksi. Pitkittäishermosäikeiden välissä on lukuisia poikittaisia yhteyksiä, ja koko hermostosta tulee tikapuumainen.
Turbellareilla ja monogeenisillä trematodeilla on aistielimiä, kuten okelleja tai silmiä. Niitä on lukuisia polyklaaveilla ja kahdesta neljään Rhabdocoeleilla ja monogeenisillä trematodeilla. Silmät ovat joko pigmenttisoluina tai pigmentti- ja aistinsoluja sisältävinä kuppeina. Kemo- ja tangoreseptoreita esiintyy laajalti, ja statokystia esiintyy Acoelassa ja muutamassa muussa muodossa.
Platyhelmintien lisääntymisjärjestelmä:
Vähäisiä Turbellaria- ja Trematoda-heimoja lukuun ottamatta platyhelminteet ovat hermafrodiitteja. Uroksen ja naaraan lisääntymiselimet ovat kussakin yksilössä erilliset ja avautuvat ulkoisesti omien huokostensa tai yhteisen sukupuoliaukon kautta. Joissakin tapauksissa gonoduktiot avautuvat ruoansulatuskanavaan, ja sukusolut vapautuvat suun kautta.
HAVAINNOT:
Joidenkin litteiden matojen kohdalla on ylimääräinen naaraspuolinen huokos tai emätinhuokos, joka palvelee kopulaation aikana. Gonoporit ovat yleensä ventraalisesti, mutta toisinaan myös dorsaalisesti. Heisimatoilla gonopori on lateraalinen.
Miehen sukuelimet koostuvat kiveksistä, joita on alkukantaisessa tilassa lukuisia ja hajallaan. Monilla kivesten määrä on vähentynyt yhteen tai kahteen, ja Acoelalta puuttuvat selvät sukurauhaset ja kanavat. Vasa efferentia vastaa kivesten lukumäärää, jos niitä on. Yleensä on pari vasa deferentiaa, jotka yhdistyvät ja avautuvat monimutkaiseksi parittelulaitteeksi.
Kopulaatioväline koostuu eversible cirrusista tai ulkonevasta peniksestä, joka on aseistettu piikeillä tai koukkupaloilla. Siihen liittyy erilaisia rauhasia. Usein on olemassa yksittäinen tai parittainen rakkula, joka toimii siemennesteen varastointisäiliönä. Naaraan sukuelimet koostuvat yhdestä tai kahdesta munasarjasta. Kun munanjohtimet ovat pareittain, ne sulautuvat distaalisesti ja muodostavat yhteisen munanjohtimen, joka avautuu parittelulaitteeseen.
Kopulaatiolaite koostuu pussista, siemennesteestä, jota kutsutaan myös siemenneste- tai kopulaatiopussiksi. Kopulaattoriin liittyy myös erilaisia rauhasia, jotka auttavat munankuorien muodostumisessa ja erittävien aineiden tuotannossa. Pitkä putkimainen tai haarautunut kohtu, johon kypsät munat kerääntyvät, on silmiinpistävä osa lisääntymisjärjestelmää.
Naaraspuolisten sukurauhasten erityispiirteenä on, että ne eroavat selvästi kahteen vyöhykkeeseen, varsinaiseen munasarjaan ja keltarauhasiin tai vitelliinirauhasiin.
Keltuainen ei koskaan sisälly munan rakenteeseen, kuten muilla eläimillä, vaan sitä tuotetaan aborttimunina, jotka sisällytetään munakapselin tai -kuoren sisälle tarjoamaan ravintoa kehittyvälle alkiolle. Cestodeissa lisääntymiselimet toistuvat jokaisessa proglottidissa. Nuorilla proglottideilla elimet ovat alkeellisia, kun taas gravidilla ne ovat pitkälle kehittyneitä.
Platyhelmintien kehitys:
VAIHTOEHDOT:
Vaikka hermafrodiittinen, ristiinhedelmöityskäytäntö on sääntö. Hedelmöitys on sisäistä ja hedelmöittyneet, kuorelliset, alkioita sisältävät munat irtoavat ulospäin. Acoelalta ja polyklaavilta puuttuvat keltarauhaset ja kehitys on suoraa. Polycladeissa syntyy vapaasti uiva toukka, jonka oletetaan ennakoivan trokoforia.
Toukkaa kutsutaan Mullerin toukaksi tai Protrochulaksi (kuva 14.29). Se on muodoltaan soikea, ja siinä on kahdeksan ulkonevaa käsivartta, joita reunustavat pitkät värekarvat, jotka muodostavat yhden yhtenäisen nauhan. Yleinen ruumiin pinta on pienten värekarvojen peitossa.
Suuaukko sijaitsee keskellä ventraalista linjaa, ja selkäpinnan etuosassa on kolme silmää. Kehityksen aikana sädekehän varret imeytyvät. Muissa ryhmissä alkionkehitys on suuresti muuttunut ja monimutkainen. 
Phylogenetic Considerations of Platyhelminthes:
Levymatojen alkuperästä ei ole yksimielistä näkemystä eläintieteilijöiden keskuudessa. Litteämatojen alkuperästä on useita näkemyksiä.
1. Lang (1881) ehdotti ctenophore- polykladiteoriaa. Tämän teorian mukaan polyklaattiset kilpikonnat ovat saattaneet kehittyä ctenoforista Platyctenean kautta. Polycladien Mullerin toukka muistuttaa jonkin verran ctenoforia.
Yhtäläiset piirteet ovat:
HAVAINNOT:
(i) Ctenoforien ja toukkien symmetrioiden samankaltaisuudet.
(ii) Mullerin toukan 8 ektodermista ciliaalista harjannetta voidaan verrata ctenoforien meridiaalisiin kampalevyihin.
(iii) Turbellarien statokystaa voidaan verrata ctenoforiin.
Tämä teoria ei saa suurta kannatusta eläintieteilijöiden keskuudessa.
2. Planulan toukka -teorian mukaan turbellariat muistuttavat jonkin verran nilviäisten planulan toukkia. Niinpä monet eläintieteilijät, kuten Hyman (1951), Jagersten (1955) ja Hadzi (1963), katsovat, että litteämadot ovat kehittyneet planulan kaltaisesta esi-isäkannasta ja että Acoela on platyhelminttien alkeellisin ryhmä (kuva 14.27A).
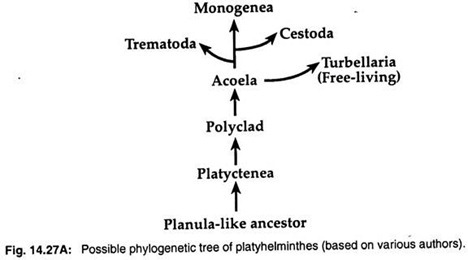
Molekulaaristen tietojen ja kladististen analyysien perusteella on ehdotettu kolmelle loisryhmälle monofyleettistä alkuperää. Kolme loisluokkaa – Trematoda, Monogenea ja Cestoda – ovat kehittyneet vapaana elävistä turbellarioista.
3. Jälleen kerran enterokoteloteorian mukaan kaikki kahdenkeskiset eläimet ovat pohjimmiltaan kotelomaattisia, ja akoelomaattiset litteät matoeläimet ovat kehittyneet kotelomaattisista esi-isistä kotelon menettämisen kautta toissijaisesti. Eläintieteilijät eivät voi hyväksyä tätä teoriaa, koska sillä on vaikea selittää joitakin näkökohtia, kuten siirtymistä kotelomaisesta tilasta akoelomaattiseen tilaan ja bilateraalisesta symmetriasta radiaaliseen symmetriaan.