Learning Outcomes
- Identify historical and potential causes of high extinction rates
Lajien määrä planeetalla tai millä tahansa maantieteellisellä alueella on tulosta kahden jatkuvasti jatkuvan evoluutioprosessin, eli eli lajinmuodostuksen ja lajikohtaisen sukupuuttoon kuolemisen, tasapainosta. Molemmat ovat makroevoluution luonnollisia ”syntymä”- ja ”kuolema”-prosesseja. Kun lajistumisnopeus alkaa ylittää sukupuutonopeuden, lajien määrä kasvaa; vastaavasti lajien määrä vähenee, kun sukupuutonopeus alkaa ylittää lajistumisnopeuden. Koko maapallon historian ajan nämä kaksi prosessia ovat vaihdelleet – joskus ne ovat johtaneet dramaattisiin muutoksiin lajien määrässä maapallolla, mikä näkyy fossiiliaineistossa (kuva 1).
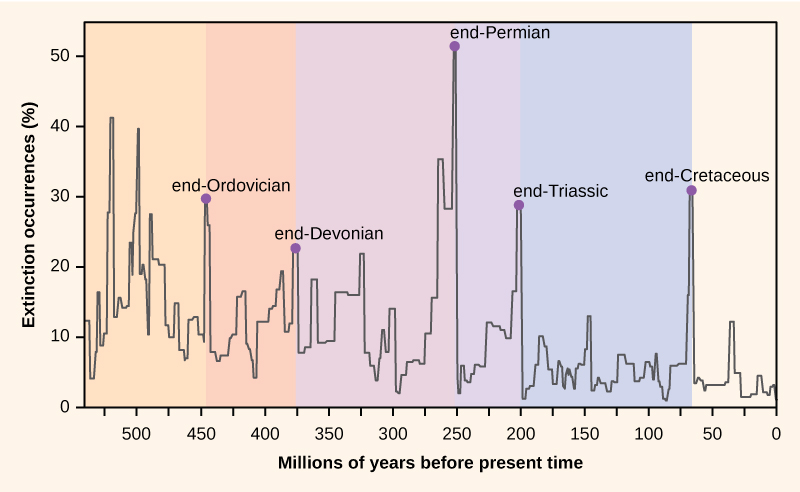
Kuva 1. Fossiiliaineistosta ilmenevät sukupuuttoprosentit ovat vaihdelleet koko maapallon historian ajan. Äkillisiä ja dramaattisia biologisen monimuotoisuuden menetyksiä, niin sanottuja joukkokuolemia, on esiintynyt viisi kertaa.
Paleontologit ovat tunnistaneet fossiiliaineistosta viisi kerrosta, joissa näyttäisi olevan havaittavissa äkillisiä ja dramaattisia (yli puolet kaikista elossa olevista lajeista katoaa fossiiliaineistosta) menetyksiä biologisessa monimuotoisuudessa. Näitä kutsutaan joukkokuolemiksi. On olemassa monia pienempiä, mutta silti dramaattisia sukupuuttoon kuolemisen tapahtumia, mutta nämä viisi massasukupuuttumista ovat herättäneet eniten tutkimusta. Voidaan väittää, että nämä viisi joukkosukupuuttoa ovat vain viisi äärimmäisintä tapahtumaa jatkuvassa suurten sukupuuttojen sarjassa, joka on jatkunut koko fanerotsooisen kauden ajan (542 miljoonaa vuotta sitten). Useimmissa tapauksissa oletetut syyt ovat edelleen kiistanalaisia; viimeisin tapahtuma näyttää kuitenkin selvältä.
Rekisteröidyt joukkosukupuuttumiset
Massasukupuuttumisten fossiiliaineisto oli perusta geologisen historian ajanjaksojen määrittelylle, joten ne tapahtuvat tyypillisesti siirtymävaiheessa geologisten ajanjaksojen välillä. Siirtyminen fossiileissa ajanjaksolta toiselle kuvastaa lajien dramaattista häviämistä ja uusien lajien asteittaista syntymistä. Nämä siirtymät näkyvät kivikerrostumissa. Taulukossa 1 on tietoja viidestä joukkosukupuutosta.
| Taulukko 1. Joukkosukupuutokset. Massasukupuutokset | ||
|---|---|---|
| Geologinen ajanjakso | Massasukupuutoksen nimi | Aika (miljoonia vuosia sitten) |
| Ordovikio-Siluria | loppu—Ordovikio O-S | 450-440 |
| Viimeinen devon | end-Devoni | 375-360 |
| Permi-Trias | Permi | 251 |
| Trias-Juraasi | -loppu-Triaskausi | 205 |
| Krea-Paleogeeni | loppukrea K-Pg (K-T) | 65.5 |
Ordoviikin ja Siluurin välinen sukupuuttotapahtuma on ensimmäinen kirjattu joukkosukupuutto ja toiseksi suurin. Tänä aikana noin 85 prosenttia merilajeista (vain harvat lajit elivät valtamerten ulkopuolella) kuoli sukupuuttoon. Päähypoteesi sen syystä on jääkausi ja sen jälkeen lämpeneminen. Itse asiassa sukupuuttotapahtuma koostuu kahdesta sukupuuttotapahtumasta, joiden välillä on noin miljoona vuotta. Ensimmäisen tapahtuman aiheutti jäähtyminen, ja toinen tapahtuma johtui sitä seuranneesta lämpenemisestä. Ilmastonmuutokset vaikuttivat lämpötiloihin ja merenpinnan tasoon. Jotkut tutkijat ovat esittäneet, että läheisen supernovan aiheuttama gammasäteilypurkaus oli mahdollinen syy Ordovikium-Silurian sukupuuttoon kuolemiseen. Gammasäteilypurkaus olisi poistanut maapallon suojaavan otsonikerroksen, jolloin auringon voimakas ultraviolettisäteily olisi päässyt maan pinnalle, ja se voisi selittää tuolloin havaitut ilmastonmuutokset. Hypoteesi on hyvin spekulatiivinen, ja maan ulkopuoliset vaikutukset maapallon historiaan ovat aktiivinen tutkimuskohde. Biologisen monimuotoisuuden elpyminen massasukupuuton jälkeen kesti paikasta riippuen 5-20 miljoonaa vuotta.
Devonin myöhäiskuopan sukupuutto saattoi tapahtua suhteellisen pitkän ajanjakson aikana. Se näyttää vaikuttaneen lähinnä merilajeihin eikä niinkään maanpäällisiä elinympäristöjä asuttaviin kasveihin tai eläimiin. Tämän sukupuuton syyt tunnetaan huonosti.
Permianin lopun sukupuutto oli elämän historian suurin. Voidaankin väittää, että maapallosta tuli lähes eloton tämän sukupuuttotapahtuman aikana. Arvioiden mukaan 96 prosenttia kaikista merilajeista ja 70 prosenttia kaikista maalla elävistä lajeista hävisi. Tänä aikana kuolivat sukupuuttoon esimerkiksi trilobiitit, ryhmä, joka selvisi Ordovikiumin ja Silurian ajan sukupuuttoon kuolemisesta. Tämän joukkosukupuuton syyt eivät ole selvillä, mutta pääepäilyksenä on laajalle levinnyt ja laajalle levinnyt tulivuoritoiminta, joka johti maapallon lämpenemiseen. Meret muuttuivat suurelta osin hapettomiksi, mikä tukahdutti meren eliöstön. Maalla elävien tetrapodien monimuotoisuuden elpyminen kesti 30 miljoonaa vuotta permikauden lopun sukupuuttoon kuolemisen jälkeen. Permin sukupuuttoon kuoleminen muutti dramaattisesti maapallon biologisen monimuotoisuuden koostumusta ja evoluution kulkua.
Trias-jurakauden sukupuuttotapahtuman syyt eivät ole selvillä, ja tutkijat esittävät hypoteeseja, joihin kuuluvat ilmastonmuutos, asteroidien isku ja tulivuorenpurkaukset. Uhanalainen sukupuuttotapahtuma tapahtui juuri ennen superkontinentti Pangaian hajoamista, vaikka viimeaikaisen tutkimuksen mukaan sukupuuttoja on saattanut tapahtua asteittaisemmin koko triaskauden ajan.
Loppukauden sukupuuttotapahtuman syyt tunnetaan parhaiten. Tämän sukupuuttotapahtuman aikana noin 65 miljoonaa vuotta sitten suurin osa dinosauruksista, jotka olivat hallitseva selkärankaisten ryhmä miljoonien vuosien ajan, katosi planeetalta (lukuun ottamatta theropodien kladeja, joka synnytti linnut).
Tämän sukupuuttoon johtaneen syyn ymmärretään nykyään olevan seurausta suuren meteoriitin eli asteroidin kataklysmisestä iskeytymisestä nykyisen Jukatanin niemimaan rannikon edustalla. Tämä hypoteesi, joka esitettiin ensimmäisen kerran vuonna 1980, oli radikaali selitys, joka perustui iridiumin (joka pääsee ilmakehään meteoreista melko tasaisella nopeudella, mutta jota ei muuten esiinny Maan pinnalla) jyrkkään piikkiin kivikerroksessa, joka merkitsi liitukauden ja paleogeenikauden välistä rajaa (kuva 2). Tämä raja merkitsi dinosaurusten ja monien muiden taksonien katoamista fossiileista. Tutkijat, jotka löysivät iridiumpiikin, tulkitsivat sen pikemminkin iridiumin nopeaksi virtaukseksi avaruudesta ilmakehään (suuren asteroidin muodossa) kuin sedimenttien laskeutumisen hidastumiseksi kyseisenä ajanjaksona. Se oli radikaali selitys, mutta raportti sopivan ikäisestä ja kokoisesta törmäyskraatterista vuonna 1991 teki hypoteesista uskottavamman. Nyt teoriaa tukevat lukuisat geologiset todisteet. Biologisen monimuotoisuuden toipumisaika liitukauden lopun sukupuuttoon kuolemisen jälkeen on geologisessa ajassa lyhyempi kuin permikauden lopun sukupuuttoon kuolemisen aikana, noin 10 miljoonan vuoden luokkaa.
Toinen mahdollisuus, joka on ehkä sattumanvarainen Jukatanin asteroidin törmäyksen kanssa, oli laajamittainen vulkaaninen liikehdintä, joka alkoi muodostua noin 66 miljoonaa vuotta sitten, eli suunnilleen samaan aikaan kuin Jukatanin asteroidin törmäys, eli liitukauden lopulla. Laavavirrat peittivät yli 50 prosenttia nykyisen Intian alueesta. Tuliperäisten kaasujen, erityisesti rikkidioksidin, vapautuminen ansojen muodostumisen aikana vaikutti osaltaan ilmastonmuutokseen, joka saattoi aiheuttaa massasukupuuton.
Harjoituskysymys

Kuva 2. Kallioperä. Iridiumkaista (luotto: USGS)
Vuonna 1980 Luis ja Walter Alvarez, Frank Asaro ja Helen Michels havaitsivat eri puolilla maailmaa piikin iridiumin pitoisuudessa sedimenttikerroksessa K-Pg-rajalla. Nämä tutkijat olettivat, että tämä iridiumpiikki johtui asteroidin törmäyksestä, joka johti K-Pg-joukkosukupuutokseen. Kuvassa 2 iridiumkerros on vaalea kaista.
Tutkijat mittasivat saniaisten itiöiden suhteellista runsautta K-Pg-rajan ylä- ja alapuolella tässä kivinäytteessä. Mikä seuraavista väittämistä edustaa todennäköisimmin heidän havaintojaan?
- Monien lajien saniaisten itiöiden runsaus havaittiin K-Pg-rajan alapuolella, mutta yhtään ei sen yläpuolella.
- Monien lajien saniaisten itiöiden runsaus havaittiin K-Pg-rajan yläpuolella, mutta yhtään ei sen alapuolella.
- Saniaisen itiöitä löytyi runsaasti sekä K-Pg-rajan ylä- että alapuolelta, mutta rajan alapuolelta löytyi vain yksi laji ja rajan yläpuolelta löytyi useita lajeja.
- Saniaisen itiöitä löytyi useista lajeista sekä rajan ylä- että alapuolelta, mutta itiöiden kokonaismäärä oli suurempi rajan alapuolella.
Pleistoseenin sukupuuttoonmeno
Pleistoseenin sukupuuttoonmeno on yksi harvinaisemmista sukupuuttoonmenoista, ja se on viimeaikainen. Tiedetään hyvin, että Pohjois-Amerikan ja jossain määrin myös Euraasian megafauna – suuret selkärankaiset eläimet – katosivat viimeisen jääkauden loppupuolella. Tämä sukupuutto näyttää tapahtuneen suhteellisen lyhyen ajanjakson aikana 10 000-12 000 vuotta sitten. Pohjois-Amerikassa menetykset olivat varsin dramaattisia, ja niihin kuuluivat muun muassa villamammutit (joiden populaatio oli olemassa noin 4 000 vuotta sitten asti eristyksissä Wrangelin saarella Kanadassa), mastodonit, jättimäiset majavat, jättiläislaiskiaiset, sapelihammaskissat ja pohjoisamerikkalainen kameli, vain muutamia mainitakseni. 1900-luvun alussa tiedemiehet esittivät ensimmäisen kerran, että ylimetsästys saattoi aiheuttaa näiden suurten eläinten nopean sukupuuttoon kuolemisen. Tämän hypoteesin tutkiminen jatkuu vielä nykyäänkin.
Yleisesti ottaen pleistoseenin sukupuuttojen ajoitus korreloi paleoihmisten saapumisen kanssa, ehkä jopa 40 000 vuotta sitten, eikä ilmastonmuutostapahtumien kanssa, mikä on tärkein kilpaileva hypoteesi näille sukupuutoille. Kuolemat alkoivat Australiassa noin 40 000-50 000 vuotta sitten, juuri sen jälkeen, kun ihmiset olivat saapuneet alueelle: pussieläinleijona, jättimäinen tonnin painoinen wombat ja useat jättiläiskengurulajit katosivat. Pohjois-Amerikassa lähes kaikkien suurten nisäkkäiden sukupuutto tapahtui 10 000-12 000 vuotta sitten. Jäljelle ovat jääneet vain pienemmät nisäkkäät, kuten karhut, hirvet, hirvet ja puumat. Lisäksi monilla syrjäisillä valtamerisaarilla monien lajien sukupuuttoon kuoleminen tapahtui samaan aikaan, kun ihmiset saapuivat alueelle. Kaikilla saarilla ei ollut suuria eläimiä, mutta kun siellä oli suuria eläimiä, ne joutuivat usein sukupuuttoon. Madagaskar asutettiin noin 2 000 vuotta sitten, ja siellä eläneet suuret nisäkkäät kuolivat sukupuuttoon. Euraasiassa ja Afrikassa ei näy tätä kaavaa, mutta niihin ei myöskään saapunut hiljattain metsästäjä-keräilijäihmisiä. Pikemminkin ihmiset saapuivat Euraasiaan satoja tuhansia vuosia sitten. Tämä aihe on edelleen aktiivisen tutkimuksen ja hypoteesien esittämisen kohteena. Vaikuttaa selvältä, että vaikka ilmastolla olisikin ollut osuutta asiaan, useimmissa tapauksissa ihmisen metsästys käynnisti sukupuuttoon kuolemisen.
Nykyaikaiset sukupuuttoon kuolemiset
Kuudes eli holoseenin aikainen joukkosukupuutto näyttää alkaneen aiemmin kuin aiemmin on uskottu, ja se johtuu suurelta osin nykyaikaisen Homo sapiens -lajin häiriötekijöistä. Holoseenikauden alusta lähtien on tapahtunut lukuisia viimeaikaisia yksittäisten lajien sukupuuttoja, jotka on kirjattu inhimillisiin kirjoituksiin. Useimmat niistä osuvat yksiin eurooppalaisten siirtomaiden laajentumisen kanssa 1500-luvulta lähtien.
Yksi varhaisemmista ja yleisesti tunnetuista esimerkeistä on dodo-lintu. Tämä outo kyyhkysen kaltainen lintu eli Mauritiuksen (saari Intian valtameressä) metsissä ja kuoli sukupuuttoon noin vuonna 1662. Merimiehet metsästivät dodoa sen lihan vuoksi, ja se oli helppo saalis, koska se lähestyi ihmisiä ilman pelkoa (dodo ei ollut kehittynyt ihmisen kanssa). Eurooppalaisten laivojen saarelle tuomat siat, rotat ja koirat tappoivat myös dodon poikasia ja munia.
Stellerin merilehmä kuoli sukupuuttoon vuonna 1768; se oli sukua manaatille ja asui luultavasti aikoinaan Pohjois-Amerikan luoteisrannikolla. Eurooppalaiset löysivät Stellerin merilehmän ensimmäisen kerran vuonna 1741, ja sitä metsästettiin liikaa lihan ja öljyn vuoksi. Viimeinen merilehmä tapettiin vuonna 1768. Näin ollen merilehmän ja eurooppalaisten ensikontaktin ja lajin sukupuuttoon kuolemisen välillä on vain 27 vuotta!
Vuoden 1900 jälkeen on kuollut sukupuuttoon useita lajeja, muun muassa seuraavat:
- Vuonna 1914 viimeinen elossa oleva matkakyyhky kuoli eläintarhassa Cincinnatissa, Ohiossa. Tämä laji oli aikoinaan pimentänyt Pohjois-Amerikan taivaan muuttomatkoillaan, mutta sitä metsästettiin liikaa ja se kärsi elinympäristönsä häviämisestä, joka johtui metsien raivaamisesta viljelysmaaksi.
- Carolinapapukaija, joka oli aikoinaan yleinen Yhdysvaltojen itäosissa, kuoli sukupuuttoon vuonna 1918. Se kärsi elinympäristönsä häviämisestä ja sitä metsästettiin, jotta se ei söisi hedelmätarhojen hedelmiä. (Papukaija söi hedelmätarhojen hedelmiä, koska sen alkuperäiset ravintokasvit hävitettiin viljelysmaan tieltä.)
- Japanin merileijona, joka asui laajalla alueella Japanin ympärillä ja Korean rannikolla, kuoli sukupuuttoon 1950-luvulla kalastajien takia.
- Karibianmeren munkkihylje oli levinnyt koko Karibianmerellä, mutta se kuoli sukupuuttoon metsästyksen vuoksi vuoteen 1952 mennessä.
Nämä ovat vain muutamia viimeisten 500 vuoden aikana kirjatuista sukupuutoista. Kansainvälinen luonnonsuojeluliitto (IUCN) pitää luetteloa sukupuuttoon kuolleista ja uhanalaisista lajeista, jota kutsutaan punaiseksi listaksi. Luettelo ei ole täydellinen, mutta siinä kuvataan 380 sukupuuttoon kuollutta selkärankaisten lajia 1500 jKr. jälkeen, joista 86 lajia on kuollut sukupuuttoon liikametsästyksen tai liikakalastuksen vuoksi.
Estimates of Present-Time Extinction Rates
Estimates of extinction rates are difficult by the fact that most extinctions are probably happening without observation. Ihmiset huomaavat todennäköisesti lintujen tai nisäkkäiden sukupuuttoon kuolemisen, varsinkin jos niitä on metsästetty tai käytetty muulla tavoin. On kuitenkin monia organismeja, jotka eivät kiinnosta ihmisiä yhtä paljon (eivät välttämättä ole vähemmän arvokkaita), ja monia, joita ei ole kuvattu.
Taustalla olevan sukupuutonopeuden on arvioitu olevan noin yksi laji miljoonaa lajia kohti vuodessa (E/MSY). Jos esimerkiksi oletetaan, että on olemassa noin kymmenen miljoonaa lajia, oletetaan, että kymmenen lajia kuolisi sukupuuttoon joka vuosi (jokainen vuosi vastaa kymmentä miljoonaa lajia vuodessa).
Eräässä nykyaikaisessa sukupuuttonopeusarviossa käytetään kirjallisiin tietoihin sisältyviä sukupuuttoja vuodesta 1500 lähtien. Pelkästään lintujen osalta tämä menetelmä antaa arvioksi 26 E/MSY. Tämä arvo voi kuitenkin olla aliarvio kolmesta syystä. Ensinnäkin monet lajit olisi kuvattu vasta paljon myöhemmin kyseisellä ajanjaksolla, joten niiden häviäminen olisi jäänyt huomaamatta. Toiseksi hiljattain sukupuuttoon kuolleiden selkärankaisten lajien määrä kasvaa, koska sukupuuttoon kuolleita lajeja kuvataan nyt luurankojäännösten perusteella. Kolmanneksi jotkin lajit ovat luultavasti jo kuolleet sukupuuttoon, vaikka luonnonsuojelijat ovatkin haluttomia nimeämään niitä sellaisiksi. Näiden tekijöiden huomioon ottaminen nostaa arvioidun sukupuutonopeuden lähemmäs 100 E/MSY:tä. Ennustettu nopeus vuosisadan loppuun mennessä on 1500 E/MSY.

Kuva 3. Tutkimukset ovat osoittaneet, että esiintyvien lajien määrä kasvaa elinympäristön koon kasvaessa. (luotto: Adam B. Smithin työn muokkaus)
Toinen lähestymistapa nykyajan sukupuuttoasteiden arviointiin on korreloida lajien häviäminen elinympäristön häviämisen kanssa mittaamalla metsien pinta-alan häviämistä ja ymmärtämällä lajien ja pinta-alan välisiä suhteita. Laji-pinta-alasuhde on nopeus, jolla uusia lajeja havaitaan, kun tutkittua pinta-alaa kasvatetaan. Tutkimukset ovat osoittaneet, että esiintyvien lajien määrä kasvaa saaren koon kasvaessa. Tämän ilmiön on osoitettu pitävän paikkansa myös muissa saarten kaltaisissa elinympäristöissä, kuten Venezuelan vuoren huipulla sijaitsevissa tepuisissa, joita ympäröi trooppinen metsä. Jos elinympäristön pinta-ala pienenee, myös siellä elävien lajien määrä vähenee. Elinympäristön häviämiseen ja lajipinta-alan suhteisiin perustuvien sukupuuttoon kuolemisen arvioiden mukaan noin 90 prosentin elinympäristön häviämisen seurauksena 50 prosenttia lajeista kuolisi sukupuuttoon. Lajipinta-ala-arviot ovat johtaneet siihen, että lajien sukupuuttoon kuolemisen määräksi on laskettu noin 1000 E/MSY ja sitä korkeammat luvut. Todelliset havainnot eivät yleensä osoita tällaista häviämismäärää, ja on esitetty, että sukupuuttoon kuoleminen on viivästynyt. Viimeaikaisessa työssä on myös kyseenalaistettu laji-pinta-ala-suhteen soveltuvuus lajien häviämistä arvioitaessa. Tässä työssä väitetään, että lajin ja pinta-alan suhde johtaa sukupuuttoon kuolemisen määrän yliarviointiin. Parempi suhde voi olla endeemisten ja pinta-alan suhde. Tämän menetelmän käyttö laskisi arviot noin 500 E/MSY:n tasolle tulevalla vuosisadalla. Huomaa, että tämä arvo on edelleen 500-kertainen tausta-arvoon verrattuna.
Kokeile
Kirjoita mukaan!
Paranna tätä sivuaOpi lisää