ADVERTISEMENTS:
I denne artikel vil vi diskutere Platyhelminthes:- 1. Levested og levested for Platyhelminthes 2. Struktur af Platyhelminthes 3. Organs of Adhesion 4. Kropsvæggen 5. Fordøjelsessystem 6. Udskillelsessystem 7. Åndedrætssystemet 8. Nervesystemet 9. Reproduktionssystem 10. Udvikling 11. Fylogenetiske overvejelser.
- Platyhelminthes’ levevis og levested:
- Struktur af Platyhelminthes:
- Hæfteorganer eller sugeorganer hos Platyhelminthes:
- Kropsvæggen hos Platyhelminthes:
- Degefærdelsessystemet hos Platyhelminthes:
- Platyhelminthes’ ekskretionssystem:
- Atmnings- og kredsløbssystemer hos Platyhelminthes:
- Nervesystem hos Platyhelminthes:
- Platyhelminthes reproduktionssystem:
- Udvikling af Platyhelminthes:
- Fylogenetiske overvejelser om Platyhelminthes:
Platyhelminthes’ levevis og levested:
Patyhelmintherne er for det meste ekto- eller endoparasitiske, og kun få er fritlevende. De fritlevende hører til klassen Turbellaria og lever i ferskvand, damme, søer, vandløb og kilder. Nogle af dem findes i kystområder i tropiske og subtropiske områder. Trematoda og cestoda er totalparasitter. I voksenstadiet parasiterer de på hvirveldyr, og i larvestadiet forekommer de som parasitter på hvirvelløse dyr.
Struktur af Platyhelminthes:
ADVERTISEMENTS:
Fylums medlemmer er normalt aflangt af udseende. Polykladerne er brede bladlignende, mens båndormene er flade og båndlignende. Kroppens kontur er generelt enkel, men nogle trematoder byder på bizzare konturer. Bændelormenes kroppe består af et antal firkantede eller rektangulære segmenter kaldet proglottider.
Den forreste proglottid er den mindste, og den bageste proglottid er den største. Det betyder, at størrelsen af proglottiden øges i anteroposterior retning. Tilstedeværelsen af proglottider giver båndormene en segmenteret tilstand. En slags pseudometamerisme forekommer hos Procerodes lobata, hvor nogle af de indre organer gentages.
Anterste og bageste ende samt dorsale og ventrale overflader er let genkendelige. Ofte er den forreste ende afgrænset fra resten af kroppen ved tilstedeværelsen af et “hoved” efterfulgt af en indsnævret “hals”.
I nogle former mangler der et decideret hoved, men den forreste ende kan registreres ved hjælp af sanseorganerne eller ved, at dens bevægelse er rettet fremad under bevægelsen. Bugfladen bærer mund- og genitalåbninger, når de er til stede.
ADVERTISER:
Størrelsen af platyhelminthes varierer fra mikroskopiske til ekstremt aflange former, der kan blive op til 10-15 meter lange (bændelorme). De fleste af medlemmerne er af små til moderate dimensioner. Platyhelminthes er generelt farveløse eller hvide i farven, fritlevende former er hvide, brune, grå eller sorte i farven. Nogle polyklader og landplanarier er med lyse farver arrangeret i mønstre.
Hæfteorganer eller sugeorganer hos Platyhelminthes:
Patyhelmintherne besidder en række forskellige adhæsions- og tilhæftningsorganer. Acetabulum eller sugeorgan i form af “sugekop” er meget almindeligt hos voksne fladorme. Hos flukes er der to sugekopper på den ventrale side af kroppen. Den ene af disse sugekopper er placeret på den forreste side af kroppen, og dens position er mere eller mindre fast.
Mens den anden, der kaldes den bageste sugekop, ikke er konstant i sin position. Hos Paramphistomum er sugekoppen bagerst i position, hos Echinostoma og Fasciola er positionen forskudt mere fortil.
I bændelorme er klæbeorganerne til stede i form af riller eller kopper og er placeret i den cephale ende. Ofte er der kroge i nærheden af disse riller eller i det evigt bughule kammer for at hjælpe med forankringen. Sugekopper forekommer også hos nogle fritlevende Planaria.
Kropsvæggen hos Platyhelminthes:
Platyhelminthes mangler exo- eller endoskelet, og som sådan er kroppen blød. Epidermis er enkeltlaget og i nogle tilfælde er den syncytial. Epidermis kan være helt eller delvist cilieret. Det subepidermale lag består af en muskulatur af cirkulære, langsgående og skråfibre. Muskelfibrene er glatte. Alle mellemrum mellem organerne er udfyldt med pakningsceller, kaldet parenkym.
De hårde dele hos Platyhelminthes er krogene og piggene. Mange tidligere arbejdere mente, at kroppen hos monogenæer, digenæer og cestoder er dækket af “kutikula”.
Nejliggørelse hos disse former giver det falske indtryk af kutikula under lysmikroskopi. Men elektronmikroskopiske undersøgelser har afsløret, at det yderste lag af kroppen er epidermis. Cellerne i dette lag indeholder mikrochondrier og forbliver sammenhængende med de underliggende celler.
Degefærdelsessystemet hos Platyhelminthes:
Digestivsystemet er fraværende hos turbellarordenen Acoela og hos cestoderne. Hos Turbellaria og Trematoda er fordøjelsessystemet repræsenteret af mund, svælg og tarm, der ender blindt, og som sådan ligner hele dispositionen af fordøjelsesorganer meget den hos anthozoer og ctenophorer.
Munden er i primitiv tilstand placeret omkring midten af buglinjen, men hos mange fladorme er placeringen forskudt fremad langs den midterste buglinje.
ADVERTISERINGER:
Munden er fraværende hos endoparasitiske rhabdocoeller. Rækken er stomodaeal af natur og er et kraftigt muskulært rør. Det viser variation i stamcellerne. Tarmen viser stor variation i form. Den kan være en simpel sæk eller kan have komplicerede forgreninger og underforgreninger. Analåbninger er sjældent til stede.
Platyhelminthes’ ekskretionssystem:
Patyhelminthes’ udskillelsessystem består hovedsageligt af langsgående vandkar eller langsgående udskillelseskanaler med et antal specialiserede udskillelsesceller, kaldet flammeceller eller flammepærer, der tilsammen kaldes protonephridier. Arrangementet af vandkarrene i kroppen giver en bred vifte af variationer.
I triclads er der to langsgående kar (kanaler), som åbner sig til ydersiden ved talrige porer (Fig. 14.28G). Rhabdocoellerne har to laterale eller et enkelt medianfartøj (Fig. 14.28E). Mediankarret åbner sig udad gennem en enkelt pore, der er placeret i den bageste ende af kroppen.
I de former, hvor der er to kar, er åbningerne enten på ventral side eller i svælget. Hos nogle former forenes de to kar i den bageste del af bagkroppen og åbner sig til ydersiden ved en enkelt medianåbning. Den primære nephridiekanal er udstrakt forgrenet, og der er accessoriske udskillelsesporer ud over nephridioporen (Fig. 14.28F).
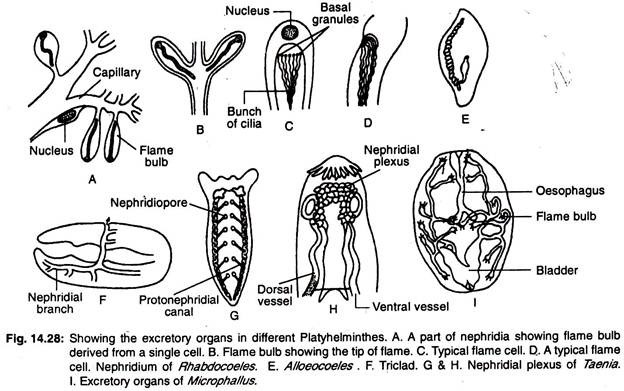
I monogene trematoder er der to langsgående udskillelseskanaler, og de to kanaler åbner hver for sig på dorsalfladen. Kanalerne udvider sig og danner udskillelsessække ved deres ender.
I andre trematoder er udskillelseskanalerne bagtil forbundet med hinanden ved en tværgående kanal og åbner sig udadtil ved en enkelt medianåbning. Den tværgående kanal dialaterer og danner i nogle tilfælde en ekskretorisk vesikel. De to langsgående kanaler kan forenes med hinanden i den bageste del og kan have en enkelt åbning.
I cestoder er der fire langsgående ekskretoriske verser. Udskillelseskarrene er forbundet af et tværgående kar, der er placeret i scolex, og danner nephridinal plexus (Fig. 14.28H) og munder ud i en kontraktile udskillelsesblære, der er placeret i den sidste proglottid.
Tallige protonephridier og et stort antal nephridioporer er til stede hos ferskvandsturbellarier, fordi tilstrømning af overdrevent vand forhindres i kroppen.
Atmnings- og kredsløbssystemer hos Platyhelminthes:
ADVERTISEMENTS:
Disse to systemer er helt fraværende i phylum. Hos nogle trematoder er tilstedeværelsen af et system af rør, kaldet lymfesystem, blevet beskrevet. Rørene anses for at have usikre funktioner.
Nervesystem hos Platyhelminthes:
Det vigtigste nervecenter eller hjerne er placeret i hovedet som et par hjerneganglier. Fra hjernen udgår flere ganglierede langsgående ledelinjer. Af disse langsgående snore bliver et par mest iøjnefaldende, og resten bliver ubetydelige. Der opstår talrige tværgående forbindelser mellem de langsgående ledbånd, og hele nervesystemet får et stigeagtigt udseende.
Sanselige organer som ocelli eller øjne er til stede hos turbellarier og monogene trematoder. De er talrige hos polyklader og to til fire hos Rhabdocoeller og monogene trematoder. Øjnene er enten til stede som pigmentceller eller i form af kopper, der indeholder pigment- og sanseceller. Chemo- og tangoreceptorer er almindeligt forekommende, og statocyster forekommer hos Acoela og få andre former.
Platyhelminthes reproduktionssystem:
Med undtagelse af nogle få Turbellaria og Trematoda er platyhelmintherne hermafroditter. Hannens og kvindens forplantningsorganer i hvert individ er adskilte og åbner sig udvendigt ved deres egne porer eller ved en fælles genitalåbning. I nogle tilfælde åbner gonodukterne sig ind i fordøjelseskanalen, og kønscellerne frigøres gennem munden.
ADVERTISER:
I nogle fladorme er der en ekstra hunporer eller vaginalporer, som tjener under parring, til stede. Gonoporerne er normalt ventral placeret, men er lejlighedsvis dorsalt placeret. Hos bændelorm er gonoporen lateralt placeret.
Hanens forplantningsorganer består af testikler, som i primitiv tilstand er talrige og spredte. Antallet af testikler er reduceret til en eller to hos mange, og hos Acoela mangler deciderede gonader og ganger. Vasa efferentia, når de er til stede, svarer til antallet af testikler, når de er til stede. I almindelighed er der et par vasa deferentia, som forenes og åbner sig i det komplicerede parringsapparat.
Kopulationsapparatet består af en evigt bevægelig cirrus eller en fremspringende penis, der er bevæbnet med pigge eller kroge. Forskellige kirtler er knyttet til den. Vesikler i enkelt eller parvis tilstand er ofte til stede og fungerer som reservoir til opbevaring af sædceller. De kvindelige forplantningsorganer består af en eller to æggestokke. De parvise æggeledere smelter sammen distalt og danner en fælles æggeleder, som munder ud i parringsapparatet.
Kopulationsapparatet består af en sæk, sædblæren eller kaldet sædslæbe eller kopulationssækken. Forskellige kirtler, som hjælper med at danne æggeskaller og producere sekretoriske stoffer, er også knyttet til parringsapparatet. En lang rørformet eller forgrenet livmoder, der har til formål at samle modne æg, udgør en iøjnefaldende del af forplantningssystemet.
Den kvindelige gonade hos platyhelminthes er ejendommelig, da den er tydeligt differentieret i to zoner, nemlig den egentlige æggestok og æggelederen eller vitellinkirtlerne.
Udblødningen er aldrig inkorporeret i ægstrukturen som hos andre dyr, men produceres som abortive æg, der indgår i ægkapslen eller -skallen for at give føde til det udviklende embryo. Hos cestoderne er forplantningsorganerne gentaget i hver proglottid. Hos unge proglottider er organerne rudimentære, mens de er højt udviklede hos de gravide proglottider.
Udvikling af Platyhelminthes:
ADVERTISEMENTS:
Og selv om de er hermafrodit, er krydsbefrugtning reglen. Befrugtningen er intern, og befrugtede, afskallede æg, der indeholder embryoner, udgydes til det ydre. Acoela og Polyclads mangler blærekirtler, og udviklingen er direkte. Hos Polyclads produceres en fritsvømmende larve, som formodes at være en forsmag på Trochophore.
Larven kaldes Muller-larve eller Protrochula (Fig. 14.29). Den er oval i formen og bærer otte fremtrædende arme, som er besat med lange cilia, der danner ét sammenhængende bånd. Den generelle kropsoverflade er dækket af små cilia.
Mundåbningen er placeret i midten af den ventrale linje, og der findes tre øjne i den forreste del af rygfladen. Under udviklingen absorberes de ciliede arme. Hos andre grupper er den embryonale udvikling stærkt modificeret og kompliceret. 
Fylogenetiske overvejelser om Platyhelminthes:
Der er ingen enstemmig opfattelse blandt zoologer med hensyn til fladormes oprindelse. Der er flere synspunkter for fladormes oprindelse.
1. Lang (1881) foreslog ctenophore- polykladeteori. Denne teori fastslår, at polykladede turbellarier kan have udviklet sig fra ctenophorer gennem Platyctenea. Mullers larve af polyklade har visse ligheder med ctenophorer.
Den ligner hinanden på følgende punkter:
ADVERTISEMENTS:
(i) Ligheder i symmetrierne mellem ctenophorerne og larven.
(ii) 8 ektodermiske cilialkamme hos Mullers larve kan sammenlignes med de meridionale kamplader hos ctenophorerne.
(iii) Statocysten hos turbellarier kan sammenlignes med ctenophorerne.
Denne teori nyder ikke megen opbakning blandt zoologer.
2. Planula-larve-teorien hævder, at turbellarierne har visse ligheder med planula-larverne hos cnidarierne. Så mange zoologer, såsom Hyman (1951), Jagersten (1955) og Hadzi (1963), mener, at fladorme har udviklet sig fra en planula-lignende stamme, og Acoela er den mest primitive gruppe blandt platyhelminter (fig. 14.27A).
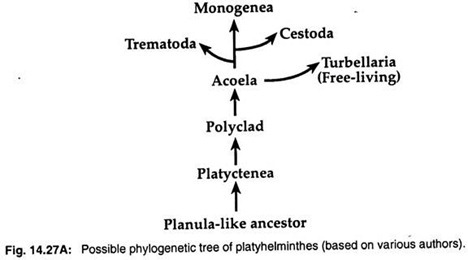
Molekylære data og kladistiske analyser er blevet foreslået en monofyletisk oprindelse for de tre parasitære grupper. De tre parasitære klasser – Trematoda, Monogenea og Cestoda – har udviklet sig fra fritlevende turbellarier.
3. Igen siger enterocoel-teorien, at alle bilaterale dyr grundlæggende er coelomate, og acoelomate fladorme har udviklet sig fra coelmate forfædre ved sekundært tab af hulrummet. Denne teori er ikke acceptabel for zoologerne, fordi den er vanskelig at forklare nogle aspekter, f.eks. ændringen fra coelomat til acoelomat tilstand og fra bilateral til radial symmetri.