ADVERTISEMENTS:
V tomto článku se budeme zabývat Platyhelminthes:- 1. Habitat a habitat Platyhelminthes 2. Živočichové. Struktura Platyhelminthes 3. Adhezivní orgány 4. Tělesná stěna 5. Trávicí soustava 6. Vylučovací soustava 7. Dýchací soustava 8. Nervová soustava 9. Reprodukční soustava 10. Vývoj 11. Fylogenetické úvahy.
- Život a prostředí Platyhelminthes:
- Struktura Platyhelminthes:
- Přilnavé nebo sací orgány u platyhelminthes:
- Tělesná stěna Platyhelminthes:
- Trávicí soustava Platyhelminthes:
- Vylučovací soustava Platyhelminthes:
- Dýchací a oběhová soustava platyhelminthes:
- Nervová soustava u Platyhelminthes:
- Reprodukční soustava Platyhelminthes:
- Vývoj Platyhelminthes:
- Fylogenetické úvahy o Platyhelminthes:
Život a prostředí Platyhelminthes:
Platýhelminty jsou většinou ekto- nebo endoparazité a jen málo z nich žije volně. Volně žijící patří do třídy Turbellaria a žijí ve sladkých vodách, rybnících, jezerech, potocích a pramenech. Některé z nich se vyskytují na pobřeží v tropických a subtropických oblastech. Trematoda a cestoda jsou totální parazité. V dospělém stadiu parazitují na obratlovcích a v larválním stadiu se vyskytují jako parazité bezobratlých živočichů.
Struktura Platyhelminthes:
PŘÍBUZNÍCI:
Příslušníci fylogeneze mají obvykle protáhlý vzhled. Mnohonožky mají tvar širokých listů, zatímco tasemnice jsou ploché a páskovité. Obrys těla je obecně jednoduchý, ale některé trematody nabízejí bizarní obrys. Tělo tasemnic je tvořeno řadou čtvercových nebo obdélníkových segmentů zvaných proglottidy.
Přední proglottida je nejmenší a nejzadnější proglottida je největší. To znamená, že velikost proglottid se zvětšuje v předozadním směru. Přítomnost proglottid propůjčuje tasemnicím segmentovaný stav. S určitým druhem pseudometamerismu se setkáváme u Procerodes lobata, u kterého se některé vnitřní orgány opakují.
Snadno rozeznatelný je přední a zadní konec, stejně jako hřbetní a břišní povrch. Přední konec je často odlišen od zbytku těla přítomností „hlavy“, za níž následuje zúžený „krk“.
U některých forem definitivní hlava chybí, ale přední konec lze rozpoznat podle smyslových orgánů nebo podle toho, že jeho pohyb směřuje dopředu během lokomoce. Ventrální povrch nese ústní a pohlavní otvory, pokud jsou přítomny.
PŘÍČINY:
Velikost platyhelmintů se pohybuje od mikroskopických až po extrémně protáhlé formy dlouhé až 10-15 metrů (tasemnice). Většina zástupců je malých až středně velkých rozměrů. Ploštěnci jsou obecně bezbarví nebo bíle zbarvení, Volně žijící formy jsou bílé, hnědé, šedé nebo černé. Někteří mnohoštětinatci a suchozemští planárníci jsou s pestrými barvami uspořádanými do obrazců.
Platýhelminthes mají různé adhezivní a přísavné orgány. Acetabulum neboli sací orgán v podobě „přísavky“ je velmi častý u dospělých ploštic. U motolic jsou na ventrální straně těla dvě přísavky. Jedna z těchto přísavek se nachází na přední straně těla a její poloha je víceméně pevná.
Zatímco druhá, nazývaná zadní přísavka, není ve své poloze stálá. U Paramphistomum je tato přísavka v zadní poloze, u Echinostoma a Fasciola je její poloha posunuta více dopředu.
U tasemnic jsou adhezivní orgány přítomny ve formě žlábků nebo pohárků a jsou umístěny na hlavovém konci. Často jsou v blízkosti těchto rýh nebo ve zvratné bukální komůrce přítomny háčky, které napomáhají ukotvení. U některých volně žijících planárií se vyskytují také přísavky.
Tělesná stěna Platyhelminthes:
Tělo Platyhelminthes postrádá exo- nebo endoskelet a jako takové je měkké. Epidermis je jednovrstevná a v některých případech je syncytiální. Epidermis může být celá nebo částečně řasnatá. Subepidermální vrstva je tvořena svalovinou z kruhových, podélných a šikmých vláken. Svalová vlákna jsou hladká. Všechny prostory mezi orgány jsou vyplněny obalovými buňkami, které se nazývají parenchym.
Tvrdé části u Platyhelminthes jsou háčky a ostny. Mnozí dřívější pracovníci se domnívali, že tělo jednobuněčných, digeneí a tasemnic je pokryto „kutikulou“.
Nepokožka u těchto forem vytváří při světelné mikroskopii falešný dojem kutikuly. Elektronmikroskopické studie však odhalily, že vnější vrstvu těla tvoří epidermis. Buňky v této vrstvě obsahují mikrochondrie a zůstávají souvislé se spodními buňkami.
Trávicí soustava Platyhelminthes:
Trávicí soustava chybí u turbelárního řádu Acoela a u cestodů. U Turbellaria a Trematoda je trávicí soustava zastoupena ústy, hltanem a střevem, které končí slepě, a tím se celé uspořádání trávicích orgánů velmi podobá uspořádání trávicí soustavy anthozoí a ctenophores.
Ústa jsou v primitivním stavu umístěna přibližně uprostřed ventrální linie, ale u mnoha plochých červů je jejich poloha posunuta dopředu podél střední ventrální linie.
ÚSTA:
U endoparazitických rhabdokolek ústa chybějí. Hltan má žaludeční charakter a je tvořen silnou svalovou trubicí. Vykazuje variabilitu v rámci fylogeneze. Střevo vykazuje velké rozdíly ve tvaru. Může mít podobu jednoduchého vaku nebo složitého větvení a podvětvení. Řitní otvory jsou přítomny jen zřídka.
Vylučovací soustava Platyhelminthes:
Vylučovací soustava platyhelminthes se skládá převážně z podélných vodních cév nebo podélných vylučovacích kanálků, které mají řadu specializovaných vylučovacích buněk, tzv. plamenných buněk nebo plamenných cibulek, souhrnně nazývaných protonefridia. Uspořádání vodních cév uvnitř těla nabízí širokou škálu variací.
U tricladů jsou dvě podélné cévy (kanálky), které se otevírají ven četnými póry (obr. 14.28G). U rhabdocoelů jsou dvě postranní nebo jedna střední céva (obr. 14.28E). Střední céva se otevírá ven jediným pórem umístěným na zadním konci těla.
U forem, kde jsou přítomny dvě cévy, jsou otvory buď na ventrální straně, nebo v hltanu. U některých forem se obě cévy spojují v zadní části a otevírají se ven jediným středním otvorem. Hlavní nefridiový kanálek je značně rozvětvený a kromě nefridioporu jsou v něm ještě další vylučovací póry (obr. 14.28F).
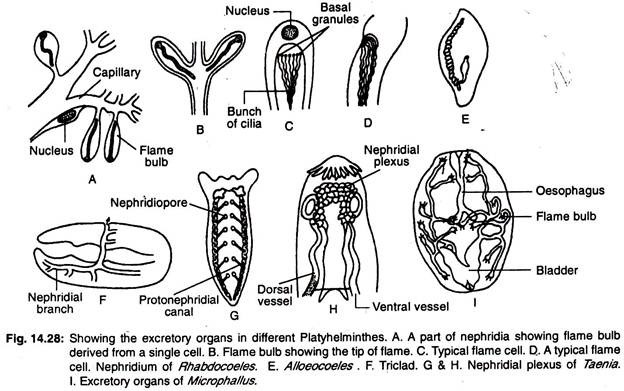
U monogenních trematod jsou dva podélné vylučovací kanálky a oba kanálky se otevírají odděleně na hřbetní ploše. Kanálky se na svých koncových koncích rozšiřují a vytvářejí vylučovací váčky.
U ostatních trematod jsou vylučovací kanálky vzájemně spojeny posteriorně příčným kanálkem a otevírají se navenek jediným středním otvorem. Příčný kanálek se vytáčí a v některých případech tvoří vylučovací měchýřek. Dva podélné kanálky se mohou vzájemně spojovat v zadní části a mohou mít jediný otvor.
U cestodů jsou čtyři podélné vylučovací kanálky. Vylučovací cévy jsou spojeny příčnou cévou umístěnou ve scolexu a tvoří nefridinální pleteň (obr. 14.28H) a otevírají se v kontraktilním vylučovacím měchýřku umístěném v poslední proglottidě.
Sladkovodní turbellariové mají četná protonefridia a velké množství nefridiopórů, protože je tak zabráněno přítoku nadměrného množství vody do těla.
Dýchací a oběhová soustava platyhelminthes:
PŘÍČINY:
Tyto dvě soustavy ve fylogenezi zcela chybí. U některých trematodů byla popsána přítomnost systému trubic, tzv. lymfatického systému. Tyto trubice jsou považovány za trubice s nejistou funkcí.
Nervová soustava u Platyhelminthes:
Hlavní nervové centrum neboli mozek se nachází v hlavě jako pár mozkových ganglií. Z mozku vychází několik gangliových podélných provazců. Z těchto podélných provazců je jeden pár nejnápadnější a zbytek se stává bezvýznamným. Mezi podélnými provazci vznikají četná příčná spojení a celá nervová soustava získává žebříkovitý vzhled.
Senzorické orgány jako ocelli nebo oči jsou přítomny u turbellarií a monogenních trematod. U mnohoštětinatců jsou početné, u rhabdocoelů a monogenních trematod jsou dva až čtyři. Oči jsou přítomny buď jako pigmentové buňky, nebo ve formě pohárků obsahujících pigmentové a smyslové buňky. Chemoreceptory a tangoreceptory jsou přítomny široce a statocysty se vyskytují u Acoela a několika dalších forem.
Reprodukční soustava Platyhelminthes:
S výjimkou několika druhů Turbellaria a Trematoda jsou platyhelminthes hermafroditi. Samčí a samičí pohlavní orgány jsou u každého jedince oddělené a navenek se otevírají vlastními póry nebo společným pohlavním otvorem. V některých případech se gonodukty otevírají do trávicího traktu a pohlavní buňky se uvolňují ústy.
PŘÍČINY:
U některých ploštic je přítomen další samičí pór nebo vaginální pór, který slouží při kopulaci. Gonopory mají obvykle ventrální polohu, ale někdy jsou i dorzální. U tasemnic je gonopor v laterální poloze.
Samčí pohlavní orgány se skládají z varlat, která jsou v primitivním stavu četná a rozptýlená. U mnohých je počet varlat redukován na jedno nebo dvě a u Acoela definitivní gonády a vývody chybějí. Vasa efferentia, pokud jsou přítomna, odpovídají počtu přítomných varlat. Obecně se vyskytuje dvojice vasa deferentia, která se spojují a ústí do složitého kopulačního aparátu.
Kopulační aparát se skládá z eversibilního cirrusu nebo z vystouplého penisu vyzbrojeného trny nebo háčky. Jsou s ním spojeny různé žlázy. Často je přítomen puchýřek v jednoduchém nebo párovém stavu, který slouží jako zásobárna pro ukládání spermií. Samičí pohlavní orgány se skládají z jednoho nebo dvou vaječníků. Párové vaječníky splývají distálně a vytvářejí společný vejcovod, který ústí do kopulačního aparátu.
Kopulační aparát se skládá z vaku, semenného váčku nebo se nazývá semenná burza či kopulační burza. S kopulačním aparátem jsou spojeny také různé žlázy, které pomáhají při tvorbě vaječných obalů a produkci sekrečních látek. Nápadnou součástí rozmnožovacího ústrojí je dlouhá trubicovitá nebo rozvětvená děloha, která slouží k hromadění zralých vajíček.
Samičí pohlavní žlázy u platyhelminthes jsou zvláštní tím, že jsou zřetelně diferencovány na dvě zóny, vlastní vaječník a žloutkové nebo vitelinové žlázy.
Žloutek není nikdy začleněn do struktury vajíčka jako u jiných živočichů, ale vzniká jako abortivní vajíčka, která se začlení do vaječné schránky nebo skořápky, aby poskytla potravu vyvíjejícímu se zárodku. U cestod se rozmnožovací orgány opakují v každé proglottidě. U mladých proglottid jsou tyto orgány rudimentární, zatímco u gravidních proglottid jsou vysoce vyvinuté.
Vývoj Platyhelminthes:
PŘÍČINY:
Ačkoli jsou hermafroditi, je praktikováno křížové oplození. Oplození je vnitřní a oplozená, skořápkovitá vajíčka obsahující zárodky se vylučují do vnějšího prostředí. Acoela a Polyclads postrádají žloutkové žlázy a vývoj je přímý. U Polycladů vzniká volně plovoucí larva, která má být předzvěstí Trochophore.
Larva se nazývá Müllerova larva nebo Protrochula (obr. 14.29). Má oválný tvar a nese osm nápadných ramen, která jsou obrostlá dlouhými řasinkami tvořícími jeden souvislý pás. Celkový povrch těla je pokryt drobnými řasinkami.
Ústní otvor se nachází uprostřed ventrální linie a v přední části hřbetní plochy jsou tři oči. Během vývoje jsou řasinková ramena pohlcena. U ostatních skupin je embryonální vývoj značně modifikovaný a komplikovaný. 
Fylogenetické úvahy o Platyhelminthes:
Mezi zoology neexistuje jednotný názor na původ ploštěnců. Existuje několik názorů na původ ploštěnců.
1. Lang (1881) navrhl teorii ctenophore- polyclad. Tato teorie uvádí, že polykládní turbanovci se mohli vyvinout z ctenophores prostřednictvím Platyctenea. Müllerova larva mnohoštětinatců vykazuje určité podobnosti s ctenophores.
Podobné znaky jsou následující:
Podobnosti:
(i) Podobnosti symetrií mezi ctenophores a larvou.
(ii) 8 ektodermálních řasnatých hřebenů Müllerovy larvy lze srovnat s meridionálními hřebínky ctenophores.
(iii) Statocystu turbellarií lze srovnat s ctenophores.
Tato teorie nemá mezi zoology velkou podporu.
2. Teorie planulární larvy tvrdí, že turbellariové mají jistou podobnost s planulární larvou hlístic. Mnozí zoologové, jako například Hyman (1951), Jagersten (1955) a Hadzi (1963), se proto domnívají, že ploštěnci se vyvinuli z předka podobného planule a Acoela je nejprimitivnější skupinou mezi plošticemi (obr. 14.27A).
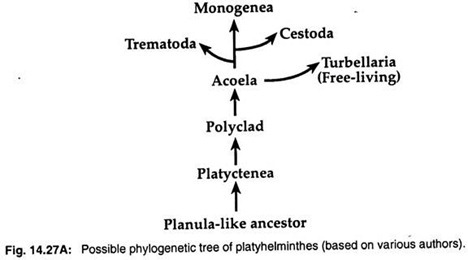
Molekulární data a kladistické analýzy naznačují monofyletický původ tří parazitických skupin. Tři parazitické třídy – Trematoda, Monogenea a Cestoda – se vyvinuly z volně žijících turbellarií.
3. Teorie enterocelií opět tvrdí, že všichni bilaterální živočichové jsou v podstatě coelomatní a acoelomatní ploštěnci se vyvinuli z coelomatních předků sekundární ztrátou dutiny. Tato teorie není pro zoology přijatelná, protože je obtížné vysvětlit některé aspekty, jako je změna z coelomatního stavu na acoelomatní a z bilaterální symetrie na radiální.