Výstupy z učení
- Identifikovat historické a potenciální příčiny vysoké míry vymírání
Počet druhů na planetě nebo v jakékoli zeměpisné oblasti je výsledkem rovnováhy dvou neustále probíhajících evolučních procesů: speciace a vymírání. Oba jsou přirozenými procesy „zrodu“ a „smrti“ makroevoluce. Když rychlost speciace začne převyšovat rychlost vymírání, počet druhů se zvýší; stejně tak se počet druhů sníží, když rychlost vymírání začne převyšovat rychlost speciace. V průběhu historie Země tyto dva procesy kolísaly – někdy vedly k dramatickým změnám v počtu druhů na Zemi, jak se odráží ve fosilním záznamu (obr. 1).
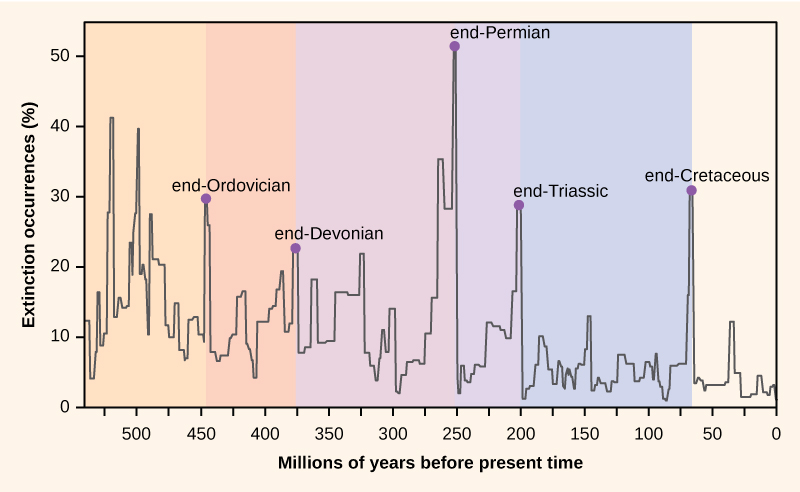
Obrázek 1. Procento výskytu vymírání, jak se odráží ve fosilním záznamu, v průběhu historie Země kolísalo. K náhlým a dramatickým ztrátám biodiverzity, tzv. masovým vymíráním, došlo pětkrát.
Paleontologové identifikovali ve fosilním záznamu pět vrstev, které zřejmě vykazují náhlé a dramatické (více než polovina všech existujících druhů mizí z fosilního záznamu) ztráty biodiverzity. Tyto případy se nazývají masová vymírání. Existuje mnoho menších, ale stále dramatických vymírání, ale těchto pět masových vymírání přitáhlo nejvíce výzkumů. Lze argumentovat, že pět masových vymírání je pouze pět nejextrémnějších událostí v nepřetržité řadě velkých vymírání v průběhu celého fanerozoika (od doby před 542 miliony let). Ve většině případů jsou hypotetické příčiny stále sporné, nicméně poslední událost se zdá být jasná.
Zaznamenaná masová vymírání
Fosilní záznam masových vymírání byl základem pro vymezení období geologické historie, takže k nim obvykle dochází na přechodu mezi geologickými obdobími. Přechod ve zkamenělinách z jednoho období do druhého odráží dramatický úbytek druhů a postupný vznik nových druhů. Tyto přechody lze pozorovat v horninových vrstvách. V tabulce 1 jsou uvedeny údaje o pěti masových vymíráních.
| Tabulka 1. Masová vymírání | ||
|---|---|---|
| Geologické období | Název masového vymírání | Čas (před miliony let) |
| Ordovik-Silur | konec-.Ordovik O-S | 450-440 |
| Pozdní devon | konec-Devon | 375-360 |
| Perm-trias | Konec permu | 251 |
| Trias-juras | konec-Trias | 205 |
| Křída-Paleogén | konec-křída K-Pg (K-T) | 65.5 |
Vymírání v ordoviku a siluru je první zaznamenané masové vymírání a druhé největší. Během tohoto období vyhynulo asi 85 % mořských druhů (jen málo druhů žilo mimo oceány). Hlavní hypotézou o jeho příčině je období zalednění a následného oteplení. Ve skutečnosti se vymírání skládá ze dvou událostí, které od sebe dělí přibližně 1 milion let. První událost byla způsobena ochlazením a druhá událost byla způsobena následným oteplením. Změny klimatu se projevily na teplotách a hladinách moří. Někteří vědci se domnívají, že možnou příčinou ordovicko-silurského vymírání byl záblesk gama záření způsobený blízkou supernovou. Záblesk gama záření mohl odstranit ochrannou ozónovou vrstvu Země a umožnit intenzivnímu ultrafialovému záření ze Slunce proniknout na zemský povrch – a to může být příčinou klimatických změn pozorovaných v té době. Tato hypotéza je velmi spekulativní a mimozemské vlivy na historii Země jsou aktivním směrem výzkumu. Obnova biologické rozmanitosti po masovém vymírání trvala v závislosti na lokalitě 5 až 20 milionů let.
K pozdně devonskému vymírání mohlo dojít za relativně dlouhou dobu. Zdá se, že postihlo především mořské druhy a ne tolik rostliny nebo živočichy obývající suchozemská stanoviště. Příčiny tohoto vymírání jsou málo známé.
Vymírání na konci permu bylo největší v historii života. Skutečně lze tvrdit, že Země se během tohoto vymírání stala téměř bez života. Podle odhadů zaniklo 96 % všech mořských druhů a 70 % všech suchozemských druhů. V této době například vyhynuli trilobiti, skupina, která přežila ordovicko-silurské vymírání. Příčiny tohoto masového vymírání nejsou jasné, ale hlavním podezřelým je rozsáhlá a rozsáhlá vulkanická činnost, která vedla k prudkému globálnímu oteplení. Oceány se staly do značné míry anoxickými, což vedlo k udušení mořského života. Po vymírání na konci permu trvalo 30 milionů let, než se obnovila rozmanitost suchozemských čtyřnožců. Permské vymírání dramaticky změnilo složení biologické rozmanitosti Země a průběh evoluce.
Příčiny triasově-jurského vymírání nejsou jasné a vědci předkládají hypotézy zahrnující klimatické změny, dopad asteroidů a sopečné erupce. K vymírání došlo těsně před rozpadem superkontinentu Pangaea, ačkoli nedávné vědecké práce naznačují, že k vymírání mohlo docházet postupněji v průběhu celého triasu.
Příčiny vymírání na konci křídy patří k těm nejlépe objasněným. Právě během tohoto vymírání před zhruba 65 miliony let zmizela z planety většina dinosaurů, po miliony let dominantní skupiny obratlovců (s výjimkou teropodního klanu, který dal vzniknout ptákům).
Příčina tohoto vymírání je dnes chápána jako důsledek kataklyzmatického dopadu velkého meteoritu neboli asteroidu u pobřeží dnešního poloostrova Yucatán. Tato hypotéza, poprvé navržená v roce 1980, byla radikálním vysvětlením založeným na prudkém nárůstu obsahu iridia (které se do naší atmosféry dostává z meteoritů poměrně konstantní rychlostí, ale jinak se na zemském povrchu nevyskytuje) v horninové vrstvě, která vyznačuje hranici mezi obdobím křídy a paleogénu (obr. 2). Tato hranice znamenala zánik dinosaurů ve zkamenělinách i mnoha dalších taxonů. Vědci, kteří iridiový hrot objevili, jej interpretovali spíše jako rychlý příliv iridia z vesmíru do atmosféry (v podobě velkého asteroidu) než jako zpomalení ukládání sedimentů v tomto období. Bylo to radikální vysvětlení, ale zpráva o nálezu impaktního kráteru odpovídajícího stáří a velikosti v roce 1991 učinila tuto hypotézu věrohodnější. Nyní tuto teorii podporuje množství geologických důkazů. Doba obnovy biodiverzity po vymírání na konci křídy je z geologického hlediska kratší než v případě vymírání na konci permu, a to řádově o 10 milionů let.
Další možností, která se možná shoduje s dopadem yucatánského asteroidu, byl rozsáhlý vulkanismus, který se začal formovat asi před 66 miliony let, tedy přibližně ve stejné době jako dopad yucatánského asteroidu, na konci křídy. Lávové proudy pokrývaly více než 50 % území dnešní Indie. Uvolňování sopečných plynů, zejména oxidu siřičitého, během vzniku pastí přispělo ke změně klimatu, která mohla vyvolat masové vymírání.
Praktická otázka

Obr. 2. Jaké jsou vlastnosti horniny pod touto vrstvou? Pás iridia (kredit: USGS)
V roce 1980 Luis a Walter Alvarezovi, Frank Asaro a Helen Michelsová objevili na celém světě nárůst koncentrace iridia v sedimentární vrstvě na hranici K-Pg. Tito badatelé vyslovili hypotézu, že tento nárůst iridia byl způsoben dopadem asteroidu, který měl za následek masové vymírání K-Pg. Na obrázku 2 je vrstva iridia světlým pásem.
Vědci měřili relativní množství výtrusů kapradin nad a pod hranicí K-Pg v tomto vzorku horniny. Které z následujících tvrzení nejpravděpodobněji vyjadřuje jejich zjištění?
- Pod hranicí K-Pg bylo nalezeno velké množství výtrusů kapradin několika druhů, ale nad ní nebyl nalezen žádný.
- Pod hranicí K-Pg bylo nalezeno velké množství výtrusů kapradin několika druhů, ale pod ní nebyl nalezen žádný.
- Hojnost výtrusů kapradin byla nalezena nad i pod hranicí K-Pg, ale pod hranicí byl nalezen pouze jeden druh a nad hranicí bylo nalezeno mnoho druhů.
- Více druhů výtrusů kapradin bylo nalezeno nad i pod hranicí, ale celkový počet výtrusů byl větší pod hranicí.
Pleistocénní vymírání
Pleistocénní vymírání patří mezi menší vymírání, a to v nedávné době. Je dobře známo, že severoamerická a do jisté míry i euroasijská megafauna – velcí obratlovci – vymizela ke konci posledního zalednění. Zdá se, že k vymírání došlo v relativně omezeném časovém období před 10 000-12 000 lety. V Severní Americe byly ztráty poměrně dramatické a týkaly se například mamutů srstnatých (jejichž zachovalá populace existovala ještě asi před 4 000 lety izolovaně na Wrangelově ostrově v Kanadě), mastodonta, obřích bobrů, obřích zemních lenochodů, šavlozubých koček a severoamerického velblouda. Na počátku 20. století vědci poprvé vyslovili domněnku, že rychlé vyhynutí těchto velkých zvířat způsobilo nadměrné lovení. Výzkum této hypotézy pokračuje dodnes.
Všeobecně lze říci, že načasování vymírání v pleistocénu souviselo s příchodem paleo-humánů, možná až před 40 000 lety, a nikoliv s klimatickými změnami, což je hlavní konkurenční hypotéza těchto vymírání. Vymírání začalo v Austrálii asi před 40 000 až 50 000 lety, těsně po příchodu člověka do oblasti: zmizel vačnatý lev, obří jednotunový vombat a několik druhů obřích klokanů. V Severní Americe došlo k vymření téměř všech velkých savců před 10 000-12 000 lety. Zůstali jen menší savci, jako jsou medvědi, losi, losi a pumy. A konečně na mnoha odlehlých oceánských ostrovech došlo k vymření mnoha druhů shodou okolností s příchodem člověka. Ne na všech ostrovech se vyskytovala velká zvířata, ale pokud se na nich vyskytovala, byla často nucena vyhynout. Madagaskar byl kolonizován asi před 2 000 lety a velcí savci, kteří tam žili, vyhynuli. Eurasie a Afrika tento vzorec nevykazují, ale také nezažily nedávný příchod lovců a sběračů. Lidé se do Eurasie dostali spíše před stovkami tisíc let. Toto téma zůstává předmětem aktivního výzkumu a hypotéz. Zdá se být jasné, že i kdyby klima hrálo určitou roli, ve většině případů urychlil vymírání lov člověka.
Vymírání v současné době
Šesté neboli holocenní masové vymírání zřejmě začalo dříve, než se dosud předpokládalo, a je z velké části způsobeno rušivou činností moderního Homo sapiens. Od počátku holocénu existuje řada nedávných vymírání jednotlivých druhů, která jsou zaznamenána v lidských spisech. Většina z nich se shoduje s expanzí evropských kolonií od roku 1500.
Jedním z dřívějších a populárně známých příkladů je pták dodo. Tento zvláštní holubovitý pták žil v lesích Mauricia (ostrov v Indickém oceánu) a vyhynul kolem roku 1662. Dodo byl loven pro své maso námořníky a byl snadnou kořistí, protože se k lidem přibližoval beze strachu (dodo se s lidmi nevyvíjel). Prasata, krysy a psi, které na ostrov přivezly evropské lodě, také zabíjeli dodova mláďata a vejce.
Morská kráva Stellerova vyhynula v roce 1768; byla příbuzná kapustňákovi a pravděpodobně kdysi žila podél severozápadního pobřeží Severní Ameriky. Mořská kráva Stellerova byla poprvé objevena Evropany v roce 1741 a byla nadměrně lovena kvůli masu a oleji. Poslední mořská kráva byla zabita v roce 1768. To činí pouhých 27 let mezi prvním kontaktem mořské krávy s Evropany a vyhynutím druhu!“
Od roku 1900 vyhynula celá řada druhů, včetně následujících:
- V roce 1914 uhynul v zoologické zahradě v Cincinnati ve státě Ohio poslední žijící holub pasažér. Tento druh kdysi při svých migracích zatemňoval oblohu Severní Ameriky, ale byl nadměrně loven a trpěl ztrátou životního prostředí, která byla důsledkem kácení lesů pro zemědělskou půdu.
- V roce 1918 vyhynul papoušek karolínský, kdysi běžný na východě Spojených států. Trpěla ztrátou biotopu a byla lovena, aby se zabránilo jejímu požírání ovoce v sadech. (Papoušek jedl ovoce ze sadů, protože jeho původní potrava byla zničena, aby uvolnila místo zemědělské půdě.)
- Morčák japonský, který obýval rozsáhlou oblast kolem Japonska a pobřeží Koreje, vyhynul v 50. letech 20. století kvůli rybářům.
- Karibský tuleň mnišský byl rozšířen po celém Karibském moři, ale v roce 1952 byl vyhuben lovem.
Toto jsou jen některé ze zaznamenaných případů vymření za posledních 500 let. Mezinárodní svaz ochrany přírody (IUCN) vede seznam vyhynulých a ohrožených druhů, který se nazývá Červený seznam. Tento seznam není úplný, ale popisuje 380 druhů obratlovců vyhynulých po roce 1500 n. l., z nichž 86 druhů vyhynulo v důsledku nadměrného lovu nebo nadměrného výlovu.
Odhady míry vymírání v současnosti
Odhadům míry vymírání brání skutečnost, že většina vymírání pravděpodobně probíhá bez pozorování. Vyhubení ptáka nebo savce si pravděpodobně všimne člověk, zejména pokud byl loven nebo jinak využíván. Existuje však mnoho organismů, které jsou pro člověka méně zajímavé (ne nutně méně hodnotné), a mnoho těch, které nejsou popsány.
Podle odhadů je rychlost vymírání přibližně jeden druh na milion druhů ročně (E/MSY). Například za předpokladu, že existuje asi deset milionů druhů, lze očekávat, že každý rok vymře deset druhů (každý rok představuje deset milionů druhů za rok).
Jeden současný odhad míry vymírání využívá vymírání v písemných záznamech od roku 1500. Jen pro ptáky dává tato metoda odhad 26 E/MSY. Tato hodnota však může být podhodnocená ze tří důvodů. Za prvé, mnoho druhů by bylo popsáno až v mnohem pozdějším časovém období, takže jejich úbytek by zůstal nepovšimnut. Za druhé, počet nedávno vyhynulých druhů obratlovců se zvyšuje, protože vyhynulé druhy jsou nyní popisovány z kosterních pozůstatků. A za třetí, některé druhy jsou pravděpodobně již vyhynulé, i když se je ochránci přírody zdráhají takto pojmenovat. Zohlednění těchto faktorů zvyšuje odhadovanou míru vymírání blíže ke 100 E/MSY. Předpokládaná míra do konce století je 1500 E/MSY.

Obrázek 3. Studie ukázaly, že počet přítomných druhů roste s velikostí stanoviště. (kredit: úprava práce Adama B. Smithe)
Druhým přístupem k odhadu míry vymírání v současnosti je korelace úbytku druhů s úbytkem stanovišť pomocí měření úbytku plochy lesa a pochopení vztahů mezi druhy a plochou. Vztah druh-plocha je rychlost, s jakou se objevují nové druhy při zvětšení sledované plochy. Studie ukázaly, že počet přítomných druhů se zvyšuje s rostoucí rozlohou ostrova. Tento jev se ukázal být platný i v jiných stanovištích podobných ostrovům, jako jsou venezuelské tepuis na vrcholcích hor, které jsou obklopeny tropickým lesem. Obrátíme-li tento vztah, zmenší-li se plocha stanoviště, sníží se i počet druhů, které zde žijí. Odhady míry vymírání založené na úbytku stanovišť a vztahu mezi rozlohou a počtem druhů naznačují, že při zhruba 90procentním úbytku stanovišť by podle očekávání vymřelo 50 % druhů. Odhady poměru plochy druhů vedly k výpočtům míry vymírání druhů na úrovni přibližně 1000 E/MSY a vyšší. Skutečná pozorování obecně nevykazují takovou míru úbytku a objevily se návrhy, že dochází ke zpoždění vymírání. Nedávné práce také zpochybnily použitelnost vztahu druh-plocha při odhadu úbytku druhů. Tato práce tvrdí, že vztah mezi druhem a plochou vede k nadhodnocení míry vymírání. Vhodnějším vztahem pro použití může být vztah endemity-oblast. Použití této metody by snížilo odhady na přibližně 500 E/MSY v nadcházejícím století. Všimněte si, že tato hodnota je stále 500násobkem míry pozadí.
Zkuste to
Přispějte!
Vylepšete tuto stránkuZjistěte více